
Глава 11. Самая универсальная болезнь — старение: роль случайных и регулярных процессов.
Никто не умирает от старости: люди в старости умирают от болезней. Да и сама старость — это болезнь, или, точнее, сумма и переплетение нарушений гомеостаза и случайных поломок.
Хотя геронтологи отнюдь не единодушны в оценке главного предмета своих исследований, но большинство их, безусловно, не считают старение болезнью. Основным их аргументом является то, что старение свойственно всем. Разве можно, рассуждают они, зачислять с определенного возраста в разряд «больных» весь род человеческий? Вот преждевременное старение, с их точки зрения, можно условно отнести к болезням — здесь они усматривают нарушение нормального процесса старения.
Между тем в свете того, что излагается в этой книге, старение является болезнью, причем болезнью не только, но своему существу, но и по определению понятия «болезнь». Рассмотрим следующий пример. По мере старения закономерно снижается скорость использования глюкозы периферическими тканями, главным образом мышечной тканью. В этом легко убедиться. В эксперименте людям разного возраста предлагается выпить определенное количество глюкозы, растворенной в воде; через некоторое время пищевая глюкоза всасывается, и концентрация ее в крови увеличивается. Чем выше возраст обследуемого, тем выше оказывается концентрация глюкозы в крови. По существу, если судить о результате пробы строго научно, то это и есть проявление сахарного диабета в точном смысле этого понятия, ибо здесь имеет место замедление использования глюкозы как топлива.
Конечно, обычное возрастное снижение использования глюкозы не есть сахарный диабет во врачебном, экспертном понимании. Но в точном смысле — это болезнь, поскольку болезнью можно назвать любое стойкое нарушение гомеостаза. (Согласно нашему более полному определению, болезнью можно назвать любое стойкое нарушение гомеостаза, ибо болезнью является любой патофизиологический процесс, увеличивающий вероятность смерти.) В данном случае, как и при сахарном диабете, проявлением нарушения гомеостаза является более длительное, чем в норме, повышение концентрации сахара в крови после глюкозной нагрузки.
Но этим примером не ограничиваются гомеостатические нарушения, присущие старению. По мере старения не только снижается скорость использования глюкозы, но и повышается выброс инсулина в ответ на глюкозу, что лежит в основе возрастного увеличения содержания жира в организме — возрастного ожирения (глава 7). В свою очередь, эти сдвиги ведут к увеличению концентрации в крови триглицеридов и холестерина (точнее, липопротеидов очень низкой и низкой плотности, в составе которых в крови находятся триглицериды и холестерин). В таблице 4 представлены данные, характеризующие эти возрастные изменения у практически здоровых мужчин.
Таблица 4. Возрастные изменения метаболических показателей у здоровых мужчин.
| Параметры | Возрастные группы (в годах) | ||||
|---|---|---|---|---|---|
| 4–19 | 20–29 | 30–39 | 40–49 | 50–59 | |
| Сахар крови натощак, ммол/л | 4,2±0,1 | 4,4±0,1 | 4,7±0,1 | 4,7±0,2 | 4,7±0,2 |
| 1 час после приема глюкозы | 4,6±0,2 | 5,5±0,2 | 6,5 ±0,6 | 7,3 ±0,5 | 7,8 ±0,4 |
| 2 часа после приема глюкозы | - | 4,7±0,3 | 5,3±0,4 | 5,8±0,5 | 6,5±0,6 |
| Инсулин крови, пмоль/л: | |||||
| натощак | 136±17,2 | 165±26,5 | 136±20,8 | 179±43,0 | 265±93,3 |
| 1 ч | 265±20,8 | 416±110,5 | 452±78,9 | 703±143,5 | 631±114,8 |
| 2 ч | - | 308±92,5 | 459±121,9 | 847±57,4 | 631±81,4 |
| Отклонение от идеальной массы тела, % | -3,4±6,3 | -8,3±2,5 | +1,2±3,0 | +3,3±3,1 | +2,3±2,4 |
| Холестерин, ммоль/л | 4,9±0,3 | 4,4±0,9 | 5,3±0,2 | 5,6±0,2 | 5,9±0,3 |
| Триглнцернды, ммоль/л | 1,06±0,1 | 1,14±0,1 | 1,36±0,1 | 1,51±0,1 | 1,71±0,04 |
В процессе старения повышается содержание в крови гонадотропинов — гормональных регуляторов репродуктивной функции, что, в свою очередь, обусловлено гипоталамическими изменениями и, в частности, снижением концентрации в гипоталамусе нейромедиаторов из группы катехоламинов — дофамина и норадреналина и т. д., и т. д. В целом по мере увеличения возраста происходят закономерные сдвиги в энергетическом, адаптационном и репродуктивном гомеостате; соответственно с той или иной степенью выраженности возникает три нормальные болезни — ожирение, гиперадаптоз и климакс. Но и этим не ограничиваются патологические процессы, органически слитые со старением. Рассмотрим еще один пример.
В старости нередко наблюдается снижение настроения, иногда приобретающее черты психической депрессии. Мы привыкли считать, что плохое настроение — это следствие усталости, телесного недомогания или в каких-то случаях — результат треволнений, накапливающихся в течение жизни. Но все не столь просто.
Психической депрессией иногда заболевают молодые телесно здоровые люди — нередко без существенного внешнего повода. Оказалось, что в этих случаях из-за обменных нарушений в гипоталамусе снижено содержание медиаторов — серотонина и норадреналина. Более того, следствием психического стресса также может быть временная апатия, поскольку в процессе антистрессорной защиты, осуществляемой гипоталамусом, в повышенном количестве расходуются медиаторы.
Но возрастное снижение концентрации этих медиаторов в гипоталамусе впрямую связано с климаксом. Если принять во внимание, что климакс как один из элементов старения развивается закономерно, то возрастное ухудшение настроения — это результат выполнения программы развития организма, т. е. еще одна «запланированная» нормальная болезнь.
В этом нет ничего неожиданного: и климакс, и психическая депрессия — это две стороны реализации закона отклонения гомеостаза, причем в обоих случаях в основе механизма реализации этого закона лежит повышение порога чувствительности гипоталамуса. На этом основана возможность диагностики психической депрессии по снижению чувствительности гипоталамуса к тормозящему эффекту «стрессорных гормонов надпочечников», например дексаметазона. Вспомним, что проба с дексаметазоном применяется и для выявления гиперадаптоза (глава 4). В этом опять-таки нет ничего необычного, т. к. болезни, сцепленные со старением, характеризуются переплетением, взаимопроникновением признаков (симптомов).
Изменения, ведущие к главным болезням старения, как бы складываются из двух частей. С одной стороны, это изменения, обусловленные гипоталамическими сдвигами, что характерно для климакса, гиперадаптоза, психической депрессии и частично — для гипертонической болезни, а также возрастного ожирения. С другой стороны, это изменения, вызванные нарушением обмена, как это имеет место при ожирении, сахарном диабете тучных, метаболической иммунодепрессии (частично при аутоиммунных болезнях), атеросклерозе и канкрофилии. Причем само развитие возрастных обменных нарушений порождается закономерными гипоталамическими сдвигами (как это отчетливо проявляется в нарушении регуляции аппетита, ведущем к ожирению).
Таким образом, не только 3 болезни, которые характеризуют изменения в трех главных гомеостатах, являются основными причинами смерти у современного человека. По существу, комплекс из 10 болезней следует рассматривать как главные болезни человека. Действительно, в этом комплексе сочетаются два способа выполнения закона отклонения гомеостаза: способ, присущий периоду беременности, формирует метаболический (обменный) компонент болезней, а способ, которым осуществляется усиление мощности главных гомеостатических систем в процессе послеэмбрионального развития и роста, создает «гипоталамическую» часть закономерных болезней и старения, наделяя само старение всеми свойствами этих болезней.
Действительно, иногда в чертах старения даже одного человека можно разглядеть все 10 главных болезней одновременно. Такой комплекс нередко проявляется при некоторых видах рака, заставляя вновь и вновь задавать себе вопрос: действительно ли существуют раздельно все эти 10 болезней или они представляют собой 10 симптомов одной интегральной болезни — старения?
Пытаясь ответить на этот вопрос, надо иметь в виду, что само старение является болезнью, или, точнее, суммой болезней гомеостаза. Соответственно не нарушение закона постоянства внутренней среды, а точное выполнение закона отклонения гомеостаза определяет болезни старения. Если эти болезни не возникают в определенный период жизни, то это указывает на отклонение от нормы.
Болезни, сцепленные со старением, начинают формироваться непосредственно вслед за окончанием развития организма, т. к. они являются продолжением развития (глава 3). Точнее, после его окончания (о чем косвенно можно судить по прекращению линейного роста тела и подготовке репродуктивной системы к процессу воспроизведения себе подобных) «движущие силы», определяющие выполнение закона отклонения гомеостаза ради развития организма, продолжают действовать, как это было и в процессе развития. Рассмотрим наиболее типичный в этом отношении пример.
В 20–25 лет вырабатывается определенное количество гонадотропинов, под воздействием которых происходит созревание детородной системы женщины. Примерно в таком возрасте организм достигает оптимального развития. Примем количество этих гормонов-регуляторов в данный период за единицу. Тогда в 45–50 лет производство регуляторных гормонов — гонадотропинов оказывается в 5 раз выше. Ясно, что в этом увеличении нет никакой физиологической потребности. Повышение продукции гонадотропинов в данной ситуации — это результат продолжающегося действия механизма, который ранее обеспечивал половое созревание. Таким механизмом является повышение порога чувствительности гипоталамуса к регулирующему действию женских половых гормонов (эстрогенов). Соответственно и явление повышенной выработки регуляторов-гонадотропинов после 25 лет — это также результат повышения порога чувствительности гипоталамуса к эстрогенным сигналам.
Вспомним и то, что в любой системе для осуществления саморегуляции необходимо сохранение количественных показателей взаимодействия между элементами. Когда взаимодействие между элементами в системах, регулируемых механизмом обратной связи, нарушается, то в результате нарушается и деятельность самой системы.
В данном примере результатом такого нарушения является прекращение способности к воспроизведению, т. е. климакс (глава 5). По механизму своего возникновения климакс представляет собой одновременно и чистое проявление процесса старения (т. к. он с неизбежной закономерностью возникает всегда), и болезни (т. к. его причиной является стойкое нарушение гомеостаза). Иными словами, климакс — это нормальная болезнь нормального старения.
Рассмотрение этого примера показывает: нет специальной программы, обусловливающей возникновение старения. Начиная с определенного возраста программа развития организма трансформируется в механизм старения и сцепленных с ним болезней. В этом смысле старение и специфические (т. е. нормальные) болезни старения являются побочным продуктом выполнения программы развития.
В свете изложенного очень, казалось бы, убедительный довод традиционных геронтологов: все то, что происходит у всех, — это норма старения, и она поэтому не должна называться болезнью, можно трактовать и совершенно противоположным образом: то, что происходит у всех, является и опасным для всех. Ведь те, кго причисляет старение к возрастной норме, молчаливо игнорируют то обстоятельство, что старение прогрессивно увеличивает риск смерти от нормальных болезней, в основе которых лежит нарушение гомеостаза. Так, между 25 и 55 годами большинство людей практически здоровы, чему и соответствует хорошее самочувствие. Но в этом возрастном интервале частота смерти от атеросклероза сосудов сердца увеличивается примерно в 100 раз.
Говоря о раннем появлении телесных признаков старения, отметим некоторые особенности, относящиеся к состоянию высшей нервной деятельности. В широко распространенное мнение о значительной гибели нервных клеток по мере увеличения возраста в настоящее время вносится очень существенное уточнение. Ускоренная гибель нервных клеток в основном происходит в тех отделах мозга, в которых в результате атеросклероза сосудов нарушается кровообращение. Иными словами, эти мозговые изменения развиваются вторично вследствие телесных изменений. Поэтому ослабление умственных способностей отнюдь не является обязательным проявлением старения. Из истории науки, искусства, философии и политики можно привести много примеров полного сохранения высокого интеллекта в глубокой старости.
Некоторые психологи вообще считают, что с годами не происходит снижения способности решать определенные задачи, характеризующие состояние интеллекта, но просто присущее обычно старению эмоциональное напряжение приводит к увеличению времени, затрачиваемого на их решение.
Весьма также примечательно, что достигнутое в наши дни значительное увеличение средней продолжительности жизни сказывается на состоянии тела, порождая современные болезни старения, тогда как вряд ли можно сомневаться в том, что человеческий разум 2000 лет назад был столь же высок, что и теперь. Гераклит, Гиппократ, Архимед, Пифагор, Эвклид, Сократ, Плутарх, Эврипид, Аристотель и многие другие умы древности были бы украшением и новейшей эпохи.
Конечно, главные болезни человека не только сцеплены с механизмом старения. Эти же болезни могут возникать случайно даже у молодых людей вследствие поломок в сложных системах физиологической регуляции организма. Более того, многие внешние факторы могут вызвать появление любой из 10 главных болезней. Поэтому в картине старения у каждого реального человека имеются свои особенности, которые зависят как от внутренних факторов старения и генетических свойств индивидуума, так и от влияния многочисленных внешних факторов, воздействию которых неизбежно и многократно подвергается каждый организм.
Мы рассмотрим все эти вопросы в специальном разделе, посвященном различным моделям возникновения болезней (глава 12). Здесь же проанализируем, как соотносится излагаемая автором модель старения с современными и с прошлыми гипотезами и теориями, трактующими сущность старения.
Прежде всего отметим; проявления старения у высших животных, включая человека, столь многообразны, что обычно интуитивно отвергается идея о наличии какой-то одной или даже ведущей причины возникновения возрастных изменений. Действительно, старение поражает каждую клетку, ткань, орган и систему. Вероятно, из-за сложности процесса старения из многих определений этого явления в научной литературе получило распространение лишь одно, основанное на соотношении старения и смертности.
В 1825 г. английский исследователь Б. Гомпертц нашел, что после 35 лет у человека показатель удельной смертности увеличивается с возрастом по экспоненциальному (логарифмическому) закону, гак что вероятность смерти примерно каждые 8 лет возрастает вдвое (рисунок 6). Соответственно старение организма определяют как естественный процесс, который по мере увеличения хронологического возраста закономерно увеличивает вероятность смерти.

Рис. 6. Данные о возрастной динамике смертности от главных болезней. По абсциссе — возраст (в годах), по ординате — смертность на 100 000 в каждом возрасте (за 1955 год, по Р. Кону):
- — общая смертность,
- — все опухоли,
- — сердечно-сосудисто — почечные;
- — атеросклероз,
- — сосудистые заболевания мозга,
- — сахарный диабет.
Исходя лишь из этого определения многие биологи пытаются понять причины старения и смерти, что вряд ли верно. Ведь в математических корреляциях типа закономерности Гомпертца отражаются изменения, происходящие в совокупности живых организмов, т. е. в популяции, а не конкретные события, лежащие в основе самого механизма старения и его проявлений. Поэтому не может быть получено какой-либо информации о сущности самого старения из анализа статистических кривых смертности (или выживания). Отсутствие достаточно разработанного определения понятия старения по его сущности не явилось, однако, препятствием к выдвижению различных гипотез о происхождении этого явления.
По мере того как возникали, совершенствовались и изменялись человеческие знания о живой природе, модифицировались и представления о старении. Особенно большое влияние на формирование взглядов в области этих сугубо биологических и человеческих проблем оказывало развитие естественных наук.
Когда великие завоевания в физике, в частности в механике, открыли перед изумленным человеком удивительный порядок в природе, где, казалось, все предопределено, или детерминировано, живая природа, включая человека, стала представляться подобием обычных, хотя и сложных машин. В связи с этим и старение начали рассматривать как естественный результат выхода из строя вследствие износа или усталости «деталей» в сложной машине человеческого тела, подобно тому как изнашиваются и устают даже совершенные металлы современных конструкций. Как отмечает В. Н. Никитин (1982), теория «изнашивания» была выдвинута Е. Мопа в 1888 г. Теория В. Н. Никитина о самозатухании синтетических процессов в геноме и в белоксинтезирующем аппарате клеток может быть отнесена в принципе к этой же группе явлений, т. к. в ней не устанавливаются причины этих возрастных изменений.
Позже были добыты знания об энергетике природы, дополнившие картину мира. Стало ясно, почему «ничто не вечно под луною»: законы термодинамики с неумолимой последовательностью возрастания энтропии несут в себе запреты, ограничивающие во времени существование любой системы. М. Рубнер (1908) выдвинул «энергетическую» теорию старения, в которой, в частности, устанавливалось, что произведение величины затраты энергии (показателя метаболизма в ккал/г/день) на максимальную продолжительность жизни (в годах) является для высших организмов постоянной величиной. В соответствии с этим правилом дается объяснение, почему животные малых размеров, у которых соответственно выше отношение поверхности к массе тела и поэтому более высоки тепловые потери, умирают быстрее.
Между тем именно живые системы организмов, благодаря обмену веществ и поступлению энергии из внешней среды, т. е. как открытые системы, обладают способностью временно противодействовать запрету, налагаемому термодинамическими законами природы (см.: Бауэр Э. С. «Теоретическая биология». Л., 1935; Шредингер Э. «Что такое жизнь? С точки зрения физика». Изд. 2-е, Л., 1972; Зотин А. И. В кн.: «Биология старения». Л., 1982).
Но если противодействие этим законам может осуществляться хотя бы в пределах видовой продолжительности жизни каждого индивидуума, то почему же все-таки его существование конечно? Очевидно, имеются какие-то специальные причины, постепенно нарушающие те свойства, которые отличают природу живых организмов от всех других явлений природы.
Когда были открыты законы генетики, а затем стали ясны структурные основы генов, построенных у высших организмов из сложных молекул дезоксирибонуклеиновых кислот (ДНК), многие ученые стали считать, что случайные или чем-либо вызванные ошибки в строении ДНК, так называемые мутации, накапливаясь, а затем постепенно воспроизводясь в процессе клеточного деления или работы клеток, ведут к старению. В этой ситуации работа организма дезорганизуется, что и соответствует физиологическому старению. К мутационным относится гипотеза Л. Сциларда (1959), связывающая старение с облучением, исходящим из космических источников; иммунологическая теория (Вернет, 1971; Уолфорд, 1969), частично — теория «катастрофы накопления ошибок» в синтезе белка (Оргел, 1963) и некоторые другие. Организм или выходит из строя вследствие накопления ошибок в своей работе, или становится уязвимым для воздействия многочисленных внешних факторов — от микробов и вирусов до эмоционального стресса, и тогда его гибель определяется суммой случайных причин, всегда существующих при взаимодействии организма со сферой его обитания1.
Мутации и накопление ошибок действительно могут иметь значение при формировании многих явлений старения. Они также играют большую роль в эволюции живой природы. Поэтому теории старения, основанные на учете этих процессов, служат и в настоящее время предметом серьезных исследований. В частности, ученых занимает вопрос, почему организмы, обладающие способностью ремонтировать ДНК при нарушении ее структуры, с течением времени делают это хуже, чем в период своего расцвета.
Теории старения «мутационного рода» в какой-то степени отвечают современным воззрениям на физическую природу мира, где многое подчиняется статистическим закономерностям, тогда как строгий порядок детерминированных явлений, характерный для классической механики, представляется слишком, грубой моделью того, что есть в живой природе2.
Особую группу представляют собой теории старения «программного» типа. Наиболее известной в этом отношении сейчас является теория, построенная на основе так называемого лимита Хайфлика, устанавливающего генетически детерминированное число возможных клеточных делений для каждого вида организмов. (В отношении этой теории имеются существенные возражения. Прежде всего высказываются сомнения в адекватности условий деления клеток в организме и вне его, где устанавливается этот лимит. В частности., нельзя исключить, что ограничение числа делений возникает иэ-за снижения чувствительности к факторам роста, что, в свою очередь, может быть обусловлено постепенным накоплением холестерина в плазматической мембране, как это происходит и в условиях организма (см. Ю. М. Лопухин и соавторы, 1983}.
К категории запрограммированной смерти (и старения), в которой предполагается наличие подобной специальной цели, ошибочно относят явления, наблюдающиеся, например, у горбуши, хотя этот вариант гибели правильнее трактовать как пример, характеризующий связь между развитием (воспроизведением) и смертью (глава 3), или как пример проявления плейотропности в действии генов (см. ниже).
Относительно большая группа теорий старения основывается на оценке деятельности организма в целом, т. е. на гак называемом системном подходе к проблеме старения.
В современную эпоху одной из ветвей такого подхода является кибернетика — наука, занимающаяся общими принципами работы и управления в различных системах и, естественно, в живых системах. В частности, здесь нелишне отметить, что конкретные данные о нейроэндокринной системе — именно о системе управления в сложных организмах — нередко служили основой для более общих выводов теоретической кибернетики, которая затем приобрела признаки интегральной науки, рассматривающей прежде всего принципы управления и взаимодействия.
Однако общие принципы кибернетики не могут путем простой экстраполяции на проблему старения объяснить его механизмы. Кибернетика помогает понять, как работает система, но она не может сама по себе, вне конкретных данных и без помощи конкретных идей объяснить, почему в процессе нормального старения системы регуляции перестают выполнять свою основную роль. На этот вопрос не дают ответа и существующие в геронтологии «системные теории» старения.
Мы кратко проследили, как общие представления о физической природе мира изменяли подходы к пониманию процесса старения. Далеко не от всего ранее приобретенного отказывались при новом витке в накоплении знаний. Однако неверно также утверждать, как это делает ряд исследователей, что существует не менее 100 различных теорий старения, или, точнее, 100 различных представлений об этом явлении. Большинство теорий, которые можно было бы найти в длинном историческом списке, постепенно утратило свое значение. Сейчас, по существу, всерьез речь может идти лишь о нескольких принципиальных подходах к проблеме старения, в частности о вероятностном и детерминированном подходах, но само содержание этих общих подходов существенно изменилось за последние годы.
Начнем с изменений, относимых к классу вероятностных событий, с которыми связывают накопление повреждений в клетках и тканях организма по мере увеличения возраста. По происхождению самих повреждающих агентов они подразделяются на агенты, исходящие из внешней окружающей среды, и агенты, порожденные внутренними закономерными процессами, происходящими в организме. По существу, только накопление повреждений, связанных с действием внешних повреждающих агентов, относится в строгом смысле к вероятностным событиям. Примером таких воздействий является етргсс, различные виды лучистой энергии, включая ультрафиолетовое облучение, химические вещества, включая канцерогены и т. д.
Несмотря на значительный реальный вклад, который могут такие факторы вносить в общую картину старения, они не являются его основной причиной. Можно, не вдаваясь в подробности, легко представить себе доводы, не позволяющие относить внешние факторы к первичным причинам старения.
Действительно, предположим, что организм находится в идеальных условиях внешней среды, которая не только не повреждает, а даже защищает его от неблагоприятных влияний. Разве в такой идеальной ситуации ие будут происходить возрастные нарушения в деятельности организма, например, такие, как выключение репродуктивной функции в определенном возрасте? Хотя внешние факторы могут ускорить наступление климакса, но отсутствие стресса не может его отменить. В то же время интерес к роли внешних факторов, которые, несомненно, могут влиять на скорость процесса старения, определяется еще также и тем, что способ действия внешних повреждающих агентов во многом совпадает с механизмом повреждений, наносимых внутренними повреждающими факторами.
Здесь, однако, необходимо внести существенное уточнение. Внутренние повреждающие события лишь в той степени относятся к вероятностным, что в отношении них никогда нельзя точно определить, «где и когда» они произойдут конкретно (например, в какой клетке или их совокупности и в какое точно время). Но само возникновение определенных внутренних причин повреждений закономерно, поскольку оно неотделимо и от свойств «материала», из которого построено «живое вещество», и от химических (биохимических) процессов, которые совершаются в организме.
Что касается «свойств материала», то хотя в живых системах постоянно происходит самообновление и хотя «атомы не стареют», но уже организованные совокупности атомов, т. е. молекулы, могут стареть за счет накопления в них повреждений. Пример такого рода — сшивка больших молекул коллагена под влиянием побочных продуктов обмена веществ, что, в частности, является одной из причин возникновения кожных морщин.
Конечно, в ходе эволюции происходило совершенствование биохимических реакций, которые, если употребить современный промышленный термин, в значительной мере приблизились к «безотходному производству», т. е. к максимально полному использованию промежуточных продуктов жизнедеятельности. Но по ряду причин, которые здесь невозможно подробно рассматривать, биохимические процессы не могут быть полностью совершенными: начиная с определенного уровня такой совершенной организации, возникала бы необходимость в значительном увеличении числа генов, функцией которых являлось бы исключительно устранение побочных реакций, а это приводило бы к нерентабельному увеличению «энергетической стоимости» поддержания самой живой системы.
Поэтому наряду с созданием необходимых защитных механизмов в процессе эволюции был «выбран» принцип плейотропного действия генов. В соответствии с этим принципом эволюционно закреплялись в генетическом коде те гены и контролируемые ими реакции (процессы), которые давали преимущества в периоде развития, роста и воспроизведения (репродукции) организмов, несмотря на возникновение в результате использования этих процессов нежелательных, в частности, повреждающих эффектов и последствий в более позднем возрасте. Но в этом периоде различные повреждения уже не сказывались на показателях воспроизведения (и поэтому не устранялись естественным отбором), а лишь вызывали явление, которое обозначается теперь словом «старение».
Наиболее яркий и существенный пример в этом отношении дает анализ явлений, связанных с приобретением в процессе эволюции способности использовать кислород в энергетических процессах. Рассмотрим в самой общей форме это явление, основываясь на трудах академика Н. М. Эмануэля, а также таких исследователей, как Д. Харман и Р. Катлер.
Зарождение жизни происходило в бескислородной среде, и появление свободного кислорода в атмосфере было обусловлено самой жизнедеятельностью. Когда концентрация кислорода в атмосфере достигла 1%, что, по ряду подсчетов, произошло примерно 1,3 млрд. лет назад, возникла токсичная среда для тех организмов, которые «научились» использовать кислород в энергетических реакциях. Но многие живые организмы сумели выжить в кислородной атмосфере, потому что у них к этому времени уже имелись защитные механизмы, противодействовавшие побочным реакциям, возникавшим при использовании кислорода, а именно — свободнорадикальным реакциям3.
В докислородную эру главной причиной, вызывающей образование свободных радикалов в организме, служила энергия Солнца, и соответственно в процессе эволюции выжили те организмы, у которых более эффективными оказались антиоксидантные (противоокислительные) механизмы. Именно эти механизмы стали использоваться для защиты от свободных радикалов, образующихся в реакциях с участием кислорода. В процессе' эволюции возник ряд антиоксидантных систем. Сейчас уже известно около 20 систем и веществ, обладающих антиоксидантнои активностью, среди которых особенно существенна роль специального фермента — супероксида дисмутазы. (Под влиянием этого фермента происходит перегруппировка двух молекул супероксида с образованием молекулярного кислорода и перекиси водорода, которая затем с помощью фермента каталазы превращается в молекулы воды.)
Побочные повреждающие эффекты любых биохимических процессов являются, по терминологии Р. Катлера, источником метаболического типа старения, и соответственно противодействующие защитные механизмы были им обозначены как антистарческие механизмы. (Вероятно, в порядке популяризации этого явления В. В. Фролькис обозначил его словом «свитаукт» — от «вита» — жизнь, «ауктум» — увеличивать: «Биология старения». Л., Наука, 1982, с. 6).
По мнению Р. Катлера, от степени эффективности антистарческих процессов при прочих равных условиях и зависит максимальная для того или иного вида продолжительность жизни, или, как эту характеристику нередко обозначают, видовая продолжительность жизни. В этом отношении представляют интерес данные, показавшие, что активность супероксида дисмутазы в тканях 12 приматов была тем выше, чем выше максимальная продолжительность жизни у изученных видов организмов. Иными словами, чем совершеннее защита от побочных реакций, возникающих при использовании кислорода, тем позднее развиваются старческие изменения, приводящие к смерти. При прочих равных условиях антнстарческие механизмы определяют видовую продолжительность жизни.
В этом отношении обратим внимание на одно очень важное обстоятельство. Когда используется термин «видовая продолжительность жизни», обычно подразумевается, что свойственная каждому виду величина этого показателя как его уникальный признак записана в генах. Между тем было замечено, что максимальная продолжительность жизни, свойственная представителям того или иного вида, находится в корреляции со скоростью гибели животных в естественной среде обитания, когда причинами гибели являются внешние факторы. Тщательнее всего изучены такого рода корреляции для птиц. Из этого делается вывод, что внутренние механизмы, определяющие гибель организма, например процессы метаболического старения и антнстарения, эволюционируют до той степени, которая определяется реальной продолжительностью жизни в естественных условиях. С этой точки зрения теперь считается правильным заменить термин «видовая продолжительность жизни» термином «максимальная продолжительность жизни».
Как уже отмечалось, ни одна система защиты не может обеспечить абсолютной защиты. Именно поэтому по мере увеличения хронологического возраста увеличивается количество и выраженность повреждений, из которых особое значение имеет повреждение генетического аппарата, и прежде всего ДНК. Хотя это явление лежит в основе возникновения мутаций — необходимого условия эволюционной изменчивости, оно должно быть предельно ограничено не только в половых, но и соматических (телесных) клетках, ибо накопление в них мутаций может приводить как к гибели клеток, так и к изменению их жизнедеятельности, например, вследствие развития аутоиммунных поражений (глава 9). Соответственно для исправления дефектов в генах существуют системы репарации (восстановления) ДНК, которые в данном контексте можно рассматривать как антистарческие системы.
Действительно, как отметили американские радиологи Р. Харт и Р. Сетлов, имеется 10-кратное различие в скорости репарации ДНК между короткоживущими и долгоживущими видами. А недавно была обнаружена более чем двукратная разница этого показателя между двумя близкими видами грызунов с различной максимальной продолжительностью жизни.
Таким образом, наряду с метаболическим процессом старения существует и функционирует процесс (и механизмы) антистарения. Оба этих процесса, если можно так выразиться, организм «несет в себе самом».
Введение понятия «антистарения» имеет еще одно очень важное значение. Как мы уже знаем, многообразие проявлений старения служит психологической преградой на пути к поиску его ключевых механизмов. Если же различные проявлении старения, например сшивка молекул коллагена и повреждение ДНК (что вторично может обусловливать самые разнообразные изменения), зависят от эффективности функционирования всего лишь двух элементов метаболического антистарення (погашения свободных радикалов и ремонта ДНК), то тогда имеются основания к изучению механизмов старения не по вторичным (внешним) его проявлениям, а по более общим и соответственно более фундаментальным процессам. Круг поиска причин старения благодаря этому сужается.
Но метаболическими явлениями не ограничиваются внутренние причины старения. Другая категория явлений относится к процессам индивидуального развития организма, т. е. к онтогенезу.
Уже исходя из чисто умозрительных и общих положений естественным представляется рассматривать старение как продолжение развития, т. е. как один из этапов онтогенеза. Но при всей самоочевидности такого подхода он очень уязвим для критики, т. к. из этого положения следует, что старение (как часть онтогенеза) так же запрограммировано, как и развитие организма. Между тем если учитывать давление естественного (эволюционного) отбора, то нет никаких оснований соглашаться с существованием программы старения (см. Введение). Поэтому идея сочетания в действии генов раннего (полезного) и позднего (повреждающего) эффектов — идея плейотропности, множественности, действия одних и тех же генов была применена не только к объяснению метаболического старения, как это рассматривалось выше, но и к старению, связанному с механизмом развития.
Основные наблюдения, которые обычно приводятся для иллюстрации этого последнего механизма, относятся к данным, показавшим, что ограничение калорийности пищевого рациона в эксперименте приводит к задержке полового созревания (т. е. к задержке развития организмов), что, в свою очередь, сочетается с замедленным старением и соответственно увеличением максимальной продолжительности жизни. Такого рода опыты, правда, тоже уязвимы для критики, поскольку ограничение калорийности пищевого рациона оказывает благоприятное действие, если оно начинается и после окончания фазы развития (в опытах на грызунах после первого года жизни). При строгом рассмотрении явлений в этих случаях не представляется, кроме того, возможным разграничить влияние снижения калорийности диеты на старение через метаболические механизмы (см. выше). Но данные, основанные на сравнении различных видов, также показывают, что чем короче у того или иного вида период развития и полового созревания, тем меньше максимальная продолжительность жизни, и, соответственно, наоборот. Так, в частности, у человека наблюдаются самый длительный по времени период полового созревания и самая значительная максимальная продолжительность жизни среди млекопитающих.
Таким образом, хотя природа связи между скоростью развития и скоростью старения еще не ясна (см. ниже), сейчас выделяется два основных механизма старения: один — метаболический и второй — связанный с механизмом развития организма4. Иными словами, старение является, по Р. Катлеру, побочным продуктом процессов обмена веществ, развития и дифферениировки в организме. Этот исследователь вводит также дополнительный компонент в данную систему, а именно — антистарческие процессы. Но этим не ограничивается схема процесса «старение — антистарение». В эту схему на основании нижеследующих соображений вводится еще один компонент.
Показатели метаболизма, старения и функции у всех млекопитающих, включая человека, в значительной степени совпадают.
Еще выше степень их однородности у высших обезьян. Например, 99% белков у человека и шимпанзе одинаковы. Вместе с тем максимальная продолжительность жизни шимпанзе в 2 раза ниже, чем у человека. Совпадают у высших организмов и вторичные признаки старения, и первичные механизмы этого явления, связанные с процессами обмена веществ и развития. Сопоставив такого рода данные, Р. Катлер пришел к выводу, что существует четвертый компонент в общей системе старения, а именно компонент «долговечности». Его рассуждения, основанные на многочисленных данных, включая археологические, ведутся на примерах, относящихся к эволюции животного мира и особенно приматов, прежде всего человека.
В процессе эволюции наблюдалось постепенное увеличение максимальной продолжительности: жизни, что удается проследить за период 60 млн. лет эволюции приматов. Весьма высокой была скорость изменения этого показателя у предков современного человека на протяжении 1,5 млн. лет, и особенно высокой (примерно 14 лет прироста за 100 000 лет) около 100 000 лет назад. Затем процесс увеличения максимальной продолжительности жизни полностью прекратился. Расчеты показывают, что за 100 000 лет быстрого роста этого показателя число полезных (адаптивных) мутаций могло равняться примерно 160–250 (при наличии у человека около 40 000 генов). Если такой небольшой процент мутаций привел к столь значительному увеличению продолжительности жизни (долговечности), то вполне естествен вывод, что эти мутации происходили в системе регуляторных генов. Иными словами, изменялась эффективность и скорость процессов, а не сама их структура за счет добавления новых свойств.
В чем заключались происходившие процессы, осталось неясным, хотя уловлены некоторые внешние признаки этих изменений. Прежде всего в ряду близких видов организмов, в частности у приматов, максимальная продолжительность жизни тем больше, чем больше размеры тела и размеры мозга, причем оба этих признака влияют независимо друг от друга. Это позволило установить простую математическую зависимость между названными параметрами, причем наблюдается хорошее совпадение между данными, предсказываемыми формулой, и наблюдаемой максимальной продолжительностью жизни (Р. Катлер, 1976).
Еще в 1965 г. Георг Сейтчер предположил, что зависимость показателя долгожительства от массы мозга можно понимать как признак какого-то системного эффекта, вероятно, как признак совершенствования гомеостаза. Обычно это предположение встречает возражение — ведь пока нет никаких свидетельств более плохой гомеостатической регуляции, например, у крысы по сравнению с обезьяной или человеком. (Это возражение нуждается в более широкой оценке. Например, хотя известно, что крысы как вид отличаются высокой приспособленностью, обеспечивающей их выживание, в самых, казалось бы, сложных (экстремальных) условиях, в эксперименте у этих животных легко возникают тяжелые нарушения при действии таких стрессов, как резкий звук или «неподвижная поза».)
Таким образом, в наиболее разработанной в отношении млекопитающих теории старения Р. Катлера можно выделить четыре компонента, образующих два блока — А и Б:
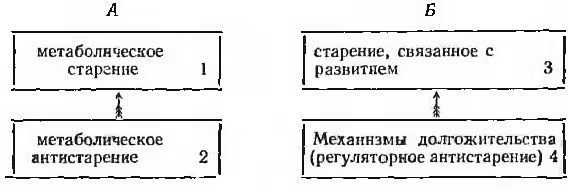
Легко заметить, что компонент Б-3 по своему наименованию соответствует онтогенетической модели старения, которая разрабатывается мною на протяжении длительного времени5. И вместе с тем то совпадение во многом лишь кажущееся.
Прежде всего, на основе онтогенетической модели рассматривается сам механизм, который определяет трансформацию (переход) программы развития в механизм старения (см. главы 4–6), тогда как в современных геронтологических концепциях (не говоря уже о более старых построениях) именно конкретные механизмы ускальзывают от рассмотрения. Во-вторых, из онтогенетической модели старения непосредственно следуют закономерности, определяющие возникновение определенных болезней, которые вследствие своей связи с механизмом развития и старения образуют группу главных (неинфекционных) болезней человека. В то же время из механизмов, рассматриваемых в современных геронтологических теориях, такая связь непосредственно не прослеживается для ряда болезней этой группы, например возрастной психической депрессии или климакса. Наконец, в-третьих, онтогенетическая модель позволяет объединить оба компонента блока Б в один компонент, т. е. единый механизм регуляторного старения и антистарення. Рассмотрим этот вопрос вначале применительно к уже сформировавшемуся виду, т. е. применительно к механизму онтогенеза, а затем и филогенеза.
Существуют значительные колебания длительности жизни у различных индивидуумов и вместе с тем закономерное логарифмическое возрастание показателя смертности по мере увеличения возраста (см. рис. 6). В свете онтогенетической модели эти колебания обусловлены различной скоростью развития, старения и болезней старения, а, в свою очередь, все эти явления в значительной степени зависят от скорости регуляторных, главным образом гипоталамических изменений. Проанализируем в качестве иллюстрации этого положения некоторые примеры (табл. 5).
Таблица 5. Механизмы естественного отбора, скорость онтогенеза, болезни старения и видовая продолжительность жизни.
| Фактор | Тип изменений | Смертность | Гипотала- мический порог | Длительность периода созревания | Скорость развития болезней старения | Видовая продол- жительность жизни |
|---|---|---|---|---|---|---|
| а. Внешние причины гибели ↑ * | филогенез | ↑ *** | ↑ | ↓ | ↑ | ↓ |
| б. Внешние причины гибели ↓ ** | филогенез | ↓ *** | ↓ | ↑ | ↓ | ↑ |
| в. Пищевой рацион ↑ | онтогенез | ↑ | ↑ | ↓ | ↑ | - |
| г. Пищевой рацион ↓ | онтогенез | ↓ | ↓ | ↑ | ↓ | - |
| д. Освещенность ↑ | онтогенез | ? | ↑ | ↓ | ↑ | - |
| е. Эстрогенный сигнал ↑ | онтогенез | ? | ↑ | ↓ | ↑ | - |
| Примечание: ↑ * — увеличение; ** — снижение; *** — от внешних факторов. | ||||||
Представим себе ситуацию, когда человек с раннего детства получает с пищей избыточное количество калорий, вследствие чего у него возникает ожирение (табл. 5, пункт «в»). Как известно, в результате этого может произойти акселерация развития (глава 14). Одним из признаков акселерации является раннее включение репродуктивной функции (аналогичным образом избыточное кормление животных ускоряет их половое созревание). Т. к. механизм полового созревания связан с повышением гипоталамического порога (глава 5), то можно сделать вывод, что в данной ситуации наблюдается увеличение скорости гипоталамических сдвигов.
Вместе с тем из онтогенетической концепции следует, что определенные явления старения (и болезней старения) являются непосредственным продолжением механизма развития. С этой точки зрения можно объяснить, почему при большей скорости гипоталамических изменений в раннем онтогенезе, т. е. при ускоренном половом созревании, должно наблюдаться ускоренное возникновение возрастной патологии, а следовательно, и меньшая продолжительность жизни. Таким образом, признак долгожительства для отдельного организма находится в зависимости от состояния систем регуляции. Далее из табл. 5 видно (пункт «г»), что противоположные изменения происходят при ограничении калорийности диеты в пререпродуктивном периоде.
Рассмотрим еще одну ситуацию (табл. 5, пункт «д»). Как известно, из экспериментов, проведенных на грызунах, избыточное освещение в пререпродуктивном периоде ускоряет половое созревание, причем экспериментально показано, что в этих условиях действительно наблюдается повышение гипоталамического порога, т. е. происходит увеличение скорости гипоталамических изменений. Соответственно и в более раннем возрасте возникают болезни, сцепленные со старением, в частности рак. В то же время избыточное освещение, примененное в периоде половозрелости, ускоряет возрастное выключение репродуктивной функции (и также ускоряет гипоталамические изменения, а значит, и повышает порог чувствительности гипоталамуса к действию половых гормонов). Следовательно, одно и то же воздействие (избыток света) может приводить как к раннему включению, так и к раннему выключению репродуктивной функции, поскольку в обоих случаях увеличивается скорость реализации одного и того же гипоталамического процесса.
Из приведенных примеров следует, что в онтогенезе у каждого в отдельности индивидуума отношение длительности пререпродуктивного периода к общей длительности жизни определяется одним и тем же регуляторным процессом, а основным элементом (компонентом) этого процесса является скорость реализации гипоталамических сдвигов (скорость выполнения закона отклонения гомеостаза, глава 3).
Теперь попытаемся оценить, применим ли этот же признак для объяснения постоянства отношения длительности пререпродуктивного периода (ПП) (или длительности периода роста) к максимальной продолжительности жизни (МПЖ) у различных видов, т. е. в филогенезе6. Для человека отношение МПЖ:ПП составляет 5:1, соответственно 100 и 20 лет. Хотя у других видов показатель не обязательно должен быть равным 5, он характеризует наличие закономерного соотношения между длительностью этих двух периодов онтогенеза; а это требует оценки данного явления в свете требований естественного отбора. Значение такой оценки заключается еще и в том, что она позволяет произвести разграничение между влиянием метаболического и регуляторного старения.
Решая задачу, учтем и различие в скорости гипоталамических изменений в онтогенетическом и филогенетическом аспектах. Для выявления роли этого признака в онтогенетическом аспекте достаточно проследить изменения на протяжении жизни одного индивидуума, например, установить связь между скоростью развития и скоростью старения в экспериментах с ограничением пищевого рациона у грызунов. Напротив, выявить роль рассматриваемого признака в филогенетическом аспекте можно только при оценке его в процессе смены поколений, что требует применения критериев естественного отбора. Теперь проанализируем две ситуации, применив для каждой из них положения как метаболического, так и онтогенетического механизма старения.
В первой ситуации имеет место увеличение гибели животных вследствие внешних причин и, следовательно, уменьшение реальной продолжительности жизни особей в рассматриваемой совокупности особей — популяции (табл. 5, пункт «а»). Т. к. естественный отбор всегда направлен на выживание вида, то в данной ситуации при отборе будут иметь преимущество особи с признаком уменьшения пререпродуктивного периода, что увеличивает вероятность оставления большего числа потомков, несмотря на уменьшение реальной продолжительности жизни. Одновременно будет наблюдаться и уменьшение общей продолжительности жизни, если эта продолжительность определяется метаболическим старением, поскольку метаболическое антистарение перестает быть селективным фактором (фактором, по которому происходит отбор, — ведь этот признак не дает преимуществ в условиях сокращения реальной длительности жизни из-за внешних причин)'. В конечном итоге в целом сохранится величина отношения МПЖ:ПП. Следовательно, в рассмотренной ситуации представление о метаболическом старении соответствует реальному положению вещей в филогенезе.
Соответствует этому критерию и онтогенетический механизм старения.
Как мы видим, здесь естественный отбор будет действовать в направлении «выбора» особей с большей скоростью повышения порога (т. к. раннее включение репродуктивной функции увеличивает вероятность оставления потомства при реально укороченной продолжительности жизни), а при большей скорости повышения порога происходит уменьшение общей продолжительности жизни (за счет более быстрого развития «нормальных болезней»), и в результате величина отношения МПЖ:ПП сохранится.
Однако анализ второй из возможных ситуаций (когда условия благоприятствуют жизни вида и в результате увеличивается реальная продолжительность жизни) на основе онтогенетической модели выявляет сохранение величины отношения МПЖ:ПП, тогда как принципы, на которых основано метаболическое старение, не соответствуют данному требованию (табл. 5, пункт «б»). Действительно, при благоприятных условиях жизни, обеспечивающих более позднее наступление гибели от внешних причин, т. е. в ситуациях, когда увеличивается реальная продолжительность жизни, метаболическое антистарение становится селективным признаком: чем позже развивается старение, тем больше вероятность оставления многочисленного потомства. Но механизм, действующий на принципах метаболических процессов, не должен приводить здесь к увеличению длительности пререпродуктивного периода — ведь и при благоприятных условиях жизни выгоднее иметь более раннее половое созревание, что обеспечивает дополнительную возможность увеличения количества потомства7. В результате отношение МПЖ:ПП должно было бы нарушиться в сторону его возрастания. Но т. к. это отношение стабильно для каждого вида, то, значит, во второй из рассматриваемых ситуаций, основываясь на явлении метаболического старения и антистарения, невозможно объяснить тенденции, контролируемые естественным отбором.
С другой стороны, этому последнему критерию соответствуют механизмы онтогенетического старения. При этом подходе применимо следующее объяснение. При благоприятных условиях, т. е. при большей реальной продолжительности жизни, преимущество получают особи с меньшей скоростью повышения гипоталамического порога — это обеспечивает большую продолжительность жизни и вероятность оставления большего количества потомков. Следовательно, этот признак становится селективным. Но он автоматически сочетается со снижением скорости повышения гипоталамичекого порога пререпродуктивного периода, т. к. оба этих параметра в целом представляют собой одно и то же явление. Поэтому хотя замедление полового созревания — отрицательный признак, раз оно уменьшает репродуктивный потенциал (неселективный признак), оно неизбежно реально вытекает из рассматриваемого регуляторного механизма старения. В результате величина отношения МПЖ:ПП сохраняется или уменьшается. Вместе с тем увеличение реальной продолжительности жизни обеспечивает в этой ситуации увеличение количества потомства, несмотря на замедление скорости полового созревания.
Имеющиеся в распоряжении науки данные показывают, что по крайней мере на протяжении последних 60 млн. лет эволюции приматов наблюдалось увеличение МПЖ. Как уже отмечалось, особенно в значительной степени этот показатель для человека возрос на протяжении предыдущих 100 000 лет (Р. Катлер, 1980). В свете этого представляет очень большой интерес; следующий вывод: отношение МПЖ:ПП, равное 5, является минимальным для животного мира. Эти данные показывают, что увеличение МПЖ сочеталось с торможением скорости развития8. Это явление, названное термином «неотения», что обозначает продленное детство, когда оно сочетается с увеличением МПЖ, соответствует селективному отбору по регуляторному (онтогенетическому), а не метаболическому типу старения.
Теперь на основе изложенного постараемся дать опенку теорий старения. Если вновь обратиться к графическому изображению наиболее современной геронтологической теории (или, по крайней мере, самой последней из них, в которой учитывается предыдущий опыт других теорий), то можно сделать следующие выводы.
Во-первых, из сравнения значения блока, А (метаболического старения) и блока Б (регуляторного старения) следует, что хотя существуют оба этих типа процессов, но регуляторное старение контролируется естественным отбором в большем числе ситуации и соответственно имеет большее эволюционное значение (т. к. эволюция у приматов пошла по линии увеличения максимальной продолжительности жизни). Соответственно в рассматриваемой схеме блок Б имеет ключевое значение.
Во-вторых, при построении блока Б нет необходимости вводить в него отдельную подсистему Б-4, т. е. механизмы долгожительства (или регуляторного антистарения). Согласно онтогенетической модели и в онтогенезе, и в филогенезе регуляторные механизмы как старения, так и антистарения осуществляются одним и тем же образом, а именно за счет замедления или ускорения одного и того же процесса (общего как для периода развития, так и старения), которым реализуется выполнение закона отклонения гомеостаза. Основной элемент этого процесса — изменение чувствительности гипоталамуса к регулирующим гомеостатическим сигналам в системах репродуктивного, энергетического и адаптационного гомеостатов.
Р. Катлер в эпиграфе к своей работе 1976 г., в которой он впервые подробно излагает свою обобщенную теорию, приводит следующее высказывание Альберта Эйнштейна:
Теория тем более впечатляюща, чем больше простота ее предпосылок, чем более различны виды предметов, к которым она относится, и более широки пределы ее применимости.
Если следовать данному критерию — наибольшей общности при наибольшей простоте, то, учитывая онтогенетический механизм, схема теории старения должна выглядеть следующим образом:
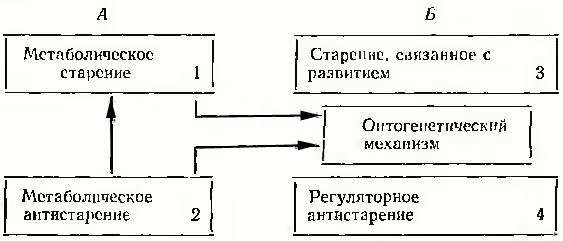
В-третьих, можно заметить, что в отличие от 4-компонентной схемы, в которой отдельно сосуществует метаболический и регуляторный блоки, в онтогенетическом механизме между блоком, А и Б имеется связь. Чтобы рассмотреть принцип такой связи, обратимся еще раз к таблице 5 (пункт «е»).
Известно, что однократное введение эстрогенов (женских половых гормонов) неполовозрелому животному в определенный период его жизни ускоряет половое созревание. Среди различных трактовок происхождения этого эффекта вполне допустима и следующая. Эстрогены усиливают синтез в гипоталамусе нейромедиаторов, прежде всего дофамина. Теперь известно, что в процессе синтеза и обмена дофамина (т. е. в процессе метаболического оборота катехоламинов) возникают свободнорадикальные реакции. Недавно было высказано предположение, что при этом происходит повреждение (или активация) клеток гипоталамического полового центра, что проявляется в повышении порога его чувствительности и тем самым в ускорении полового созревания9.
Когда под влиянием избыточного питания ускоряется половое созревание, в этом комплексном эффекте также может принимать участие аналогичный механизм — ведь чем больше генерация энергии и интенсивнее обмен веществ, тем больше «выход» свободных радикалов. С этой точки зрения можно трактовать и константу М. Рубнера (см. выше), а именно: величина расхода калорий потому связана с величиной продолжительности жизни, что энергетические процессы влияют как на метаболический, так и на регуляторный компоненты старения.
Таким образом, в рамках онтогенетической модели удается наметить существование связи между вероятностными и детерминированными механизмами развития и соответственно старения, а это является необходимым шагом на пути к интеграции в биомедицине.
Кроме того, адаптация к внешним неблагоприятным факторам, влияющим на показатель смертности в популяции, осуществляется за счет изменений чувствительности гипоталамического порога к регулирующим сигналам, что во многом определяет различную уязвимость в отношении стресса и болезней, наблюдаемую у молодых и старых животных. Рассмотрим известный пример из работ Дж. Христиана (1976).
Когда плотность популяции, например за счет увеличения рождаемости, достигает определенной величины, у животных происходит повышение секреции главных стрессовых горлеонов — кортикотропина и кортикостерона. С одной стороны, эти гормоны снижают активность иммунитета и тем самым увеличивают смертность от инфекций и паразитарных болезней, а с другой — путем действия АКТГ на гипоталамический половой центр тормозится репродуктивная функция. При этом степень и длительность повышения уровня стрессорных гормонов, как и степень чувствительности полового центра к подавлению, определяются возрастом особей, подвергающихся популяционному стрессу. У старых животных из-за резистентности к подавлению, что лежит в основе гиперадаптоза (глава 4), дольше длится стрессорное состояние, а у молодых животных из-за большей чувствительности к подавлению короче, чем у старых, продолжительность стресса, но значительнее подавляется способность к размножению. В конечном итоге старые — больше умирают, молодые — меньше размножаются. В результате восстанавливается оптимальный объем популяции, причем сохраняется возможность вступления молодых животных в цикл воспроизведения при возникновении благоприятных условий. Легко увидеть, что метаболические процессы старения и антистарения не обеспечивают подобных возможностей адаптации.
Таким образом, исходя из представления об онтогенетическом механизме развития и старения, закономерно сделать следующий вывод. Единой мерой, определяющей как скорость регуляторного старения у отдельного индивидуума, так и различия в максимальной продолжительности жизни у отдельных видов с близкими физиологическими характеристиками (например, у млекопитающих), является скорость выполнения закона отклонения гомеостаза или, что почти одно и то же, скорость гипоталамических изменений.
С этой точки зрения может быть предложен решающий эксперимент для оценки степени правдоподобности филогенетического аспекта онтогенетической модели. Этот эксперимент, если модель верна, должен показать наличие обратной зависимости между максимальной продолжительностью жизни вида и скоростью изменения гипоталамического порога. Иными словами, чем выше эта скорость, тем меньше должна быть максимальная продолжительность жизни в ряду близких видов.
Когда речь идет об онтогенетической модели применительно к проблеме старения, то это, по существу, не должно выглядеть чем-то необычным. Многие исследователи исходя из общих соображений представляли старение как часть онтогенеза, и новизна онтогенетической модели в этом отношении обусловлена установлением гипоталамического механизма взаимосвязи между развитием и старением. Но то обстоятельство, что побочным продуктом программы развития является не только старение, но и определенные болезни, а именно — главные болезни человека (и высших животных), уже никак не воспринимается в рамках традиционной геронтологии.
Но чтобы разобраться, почему существуют столь противоположные мнения о связи старения и определенной группы болезни необходимо ответить на более общий вопрос: а что вообще является причиной болезней?
Об этом — в следующей главе.
1 Подобные соображения о роли внешних факторов в причинах смерти рассматриваются в метаболических теориях (см.: Б. Стрелер. «Время, клетки, старение». М., 1964) и в «стрессорной теории старения» Г. Селье (1976).
2 К вероятностным теориям относится генно-регуляторная концепция В. В. Фролькиса (1970, 1975), т. к. первичные изменения, постулируемые ею, вызываются функциональными поломками в системе регуляторных генов, а причины поломок не уточняются.
3 Свободными радикалами называют атомы или молекулы, имеющие неспаренный электрон. Этот неспареиный электрон делает их чрезвычайно реакционноспособными. Свободные радикалы могут возникать в клетке по многим механизмам, но чаще и интенсивнее всего — в качестве нестойких промежуточных продуктов нормального обмена веществ, например при окислительных процессах, в реакциях, связанных с образованием АТФ (адеиозинтрифосфата), играющего роль связующего звена между процессами, поставляющими энергию, и процессами, в которых происходит ее использование. Свободные радикалы часто вызывают перекисное окисление ненасыщенных жирных кислот, входящих в структуры мембран клетки, что приводит к их нарушению и соответственно нарушению функции клетки, в частности, в результате повреждения мембран лизосом (где находятся «расщепляющие ферменты») и мембран митохондрий (где происходят окислительные реакции энергетических субстратов). Накопление старческого пигмента — липофусцина также связано с перекисным окислением лепидов и некоторых других соединений. Особенно важно, что свободные радикалы могут взаимодействовать с ДНК и белками, а это может вызвать как мутации, так и другие повреждения генома, например депреессито «блокированных» генов (см. глава 10) или нарушение дифференцировки клетки (ее деднфференцнровку), что и характерно для старения.
4 Подчеркнем еще раз преимущество такого подхода, позволяющего не рассматривать как отдельные механизмы старения (и соответственно в качестве отдельных теорий старения) такие явления, как сшивку макромолекул белка или ДНК; соматические мутации, в частности в клетках иммунной системы; катастрофу накопления ошибок в синтезе белков (ферментов); гибель клеток вследствие повреждений; свободнорадикальные реакции или депуринизацию ДНК и т. д.
5 Автор считает необходимым отвести возможные подозрения в заимствовании. Хотя термин «онтогенетическая модель» старения и сцепленных с ним болезней стал употребляться в печати лишь с 1983 г. в связи с предложением редактора моей книги «Закон отклонения гомеостаза и болезни старения» (Бостон, 1981) о разработке в ней новой модели медицины (что, естественно, потребовало дать наименование модели, см. глава 12), сам регуляторный принцип перехода программы развития организма в механизм старения был опубликован мною в зарубежной печати в 1971 г. (Lancet, 1971, № 1. p. 1211–1219), а в отечественной литературе — начиная с 1958 года. В то же время, Р. Катлер впервые высказал рассматриваемые здесь идеи в 1972 г., а в терминологии, указывающей на связь старения с развитием, лишь в 1976–1978 гг.
6 Филогенез — эволюционная история (зависимость) развития видов.
7Здесь не рассматриваются более сложные механизмы, в которых допускается, что метаболическое старение (и соответственно антистарение) влияет на скорость реализации регуляторного (онтогенетического) старения (см. ниже).
8 Отмечают, что человек, например, на всех стадиях своего развития имеет больше черт «детскости», чем близкий ему вид — шимпанзе, у которого МПЖ в 2 раза меньше, чем у человека.
9 В пользу реальности подобного влияния, при котором само выполнение функции вызывает отклонение гомеостаза, свидетельствуют данные, показавшие, что применение эстрогенов у девочек с врожденным отсутствием яичников вызывает повышение порога чувствительности гипоталамуса к эстрогенам. Это явление укладывается также в «каскадный механизм старения», предложенный американским нейрофизиологом К. Финчем (1976), причем, как было показано этим исследователем п соавторами в дальнейшем (1983), устранение эстрогенов у животных (в эксперименте) замедляет развитие возрастных гипоталамическнх сдвигов.