
Опухоли, гены и эволюция

Андрей Петрович Козлов доктор биологических наук, Биомедицинский центр, Санкт-Петербург «Химия и жизнь» №8, 2009
В 1972 году, после окончания Ленинградского государственного университета, поступив в аспирантуру НИИ онкологии, я прочел «Руководство по общей онкологии», изданное под редакцией выдающегося ученого Н. Н. Петрова. Одна из глав этой книги была посвящена сравнительной онкологии. Я помню, как был поражен тем, насколько широко распространены опухоли. В университете, славящемся своей эволюционной школой, нам об этом не рассказывали.
А. П. Козлов является создателем нескольких лабораторий и многих успешных исследовательских программ. Например, он участвовал в разработке российской федеральной программы «Вакцины нового поколения и медицинские диагностические системы будущего», финансировавшегося министерством науки России в 1997–2002 гг.
Опухоли повсюду
В 1802 году одно из научных обществ Эдинбурга впервые сформулировало вопрос: «Встречаются ли у примитивных организмов заболевания, напоминающие рак у человека?» После 1838 года, когда немецкий физиолог Иоганн Мюллер установил, что опухоли человека состоят из клеток, основным инструментом в повседневной клинической диагностике злокачественных новообразований стала микроскопия. До конца XIX века исследователи микроскопически идентифицировали опухоли у домашних животных, рыб и моллюсков, то есть в первую очередь у организмов, имеющих хозяйственное значение для человека.
В XX веке опухоли или опухолеподобные процессы нашли практически у всех многоклеточных организмов, в том числе у беспозвоночных, и даже у гадрозавров, обитавших в меловом периоде. (У других семейств динозавров опухолей пока не обнаружили. Возможно, различные группы динозавров имели разную предрасположенность к новообразованиям.) Вообще, чем выше организация животных, тем чаще у них возникают опухоли. Например, у насекомых они встречаются чаще, чем у всех более низкоорганизованных беспозвоночных, у позвоночных чаще, чем у низших хордовых (оболочников и ланцетников), а у костистых рыб чаще, чем у хрящевых. Опухоли беспозвоночных и низших позвоночных отличаются от опухолей млекопитающих. Так, новообразования рыб, как правило, менее агрессивны, более дифференцированны и реже метастазируют. У рыб и тритонов описаны сезонные опухоли с полной регрессией — феномен, неизвестный у млекопитающих.

Шапочка-опухоль золотой рыбки. Изображение: «Химия и жизнь»
Опухоли не редкость и у растений. Их вызывают самые разные факторы внешней среды (например, облучение или ранения) или патогены (бактерии, вирусы, простейшие). Большую группу опухолей растений составляют спонтанные новообразования генетической природы.
Примером растительных опухолей могут служить так называемые корончатые галлы, возникающие под влиянием опухолеобразующих бактерий Agrobacterium tumefaciens. Исследования последних десятилетий показали, что A. tumefaciens трансформирует клетки растений фрагментом ДНК (T-DNA, transferred DNA) большой Ti-плазмиды (tumor inducing plasmid). Т-ДНК включает растительные онкогены, продукты которых, ауксины и цитокины, стимулируют деление клеток и образование опухоли. Кроме того, Т-ДНК содержит ген, кодирующий фермент синтеза опинов — производных аминокислот, которые бактерии (но не растение) используют в качестве источника углерода, азота и энергии. Сильно пораженные растения становятся чахлыми, мелкими и непродуктивными. Таким образом, корончатые галлы представляют собой настоящую инфекционную опухолевую патологию у растений и по своей природе близки к истинным опухолям животных.
Систематизировав и обобщив множество наблюдений и экспериментальных фактов, сравнительная онкология сформулировала представление о широкой распространенности опухолей у многоклеточных организмов, включая беспозвоночных и растения. Поэтому опухоли вполне могут играть какую-то роль в эволюции многоклеточных.
В этой статье мы рассмотрим свидетельства в пользу возможной положительной роли опухолей в эволюции, в том числе наши собственные экспериментальные данные, полученные в Биомедицинском центре. При этом вслед за Н. Н. Петровым мы будем рассматривать не отдельные формы злокачественных опухолей, а все виды избыточного клеточного роста у многоклеточных организмов, включая опухолеподобные процессы у беспозвоночных и растений, доброкачественные опухоли и опухоли на ранних стадиях прогрессии.
Опухоли — эволюции
Многие опухоли и полученные из них клеточные линии продуцируют белки, нехарактерные для тканей или клеточных типов, из которых произошли эти опухоли.
В конце 1980-х — начале 1990-х годов мы попробовали определить максимальное число генов, которые экспрессируются в нормальных клетках организма и только в опухолях. (Эксперименты выполнены в сотрудничестве с доктором биологических наук В. И. Евтушенко и профессором К. П. Хансоном.) В этих работах мы использовали насыщающую молекулярную гибридизацию смеси РНК, выделенной из 12 органов крысы и из эмбрионов крысы на трех стадиях развития («суммарная РНК»). Зондом служила неповторяющаяся ДНК из печени крыс, меченная радиоактивным йодом. (С неповторяющейся ДНК связано большинство структурных генов.)
В результате экспериментов, схема которых приведена на рисунке, мы определили, что в нормальной клетке экспрессируется примерно 55–56% генома, а в опухолевых клетках дополнительно работают несколько сотен или тысяч генов, которые «молчат» в нормальных тканях, в том числе эмбриональных. Следовательно, в клетках опухоли идет активный синтез белков и нуклеиновых кислот, что могло сыграть важную роль в эволюции.
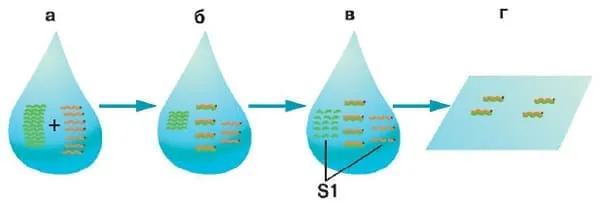
Вот так делают насыщающую молекулярную гибридизацию:
а — в растворе смешивают избыточное количество РНК опухолевой или нормальной тканей и фрагменты радиоактивно меченной ДНК (с точкой); б — при длительной инкубации гомологичные РНК и ДНК связываются друг с другом, образуя гибридные двухцепочечные молекулы (дуплексы); в — РНК и ДНК, которые не образовали дуплексов, разрушают нуклеазой S1, расщепляющей только однонитевые нуклеиновые кислоты; г — дуплексы ДНК-РНК сорбируют на специальных фильтрах, после чего по соотношению исходного количества метки и метки, оставшейся на фильтре, оценивают, какая часть генома транскрибируется. Изображение: «Химия и жизнь»
Другое эволюционно важное свойство опухолевых клеток — их способность к дифференцировке с одновременной потерей злокачественности.
В 70-х годах XX века ученые из разных стран установили, что диметилсульфоксид, добавленный в культуру опухолевых клеток, вызывает их дифференцировку. Аналогично действует на некоторые мышиные миелоидные клеточные линии фактор CSF, вызывающий образование колоний. В 1975 году Беатриса Минц (США) и Карл Ильмензее (Швейцария) установили, что клетки тератокарциномы, введенные в мышиный эмбрион на стадии бластулы, утрачивают злокачественность и участвуют в образовании нормальных тканей. В результате получаются нормальные генетически мозаичные мыши. Таким образом, даже клетки опухолей млекопитающих способны к дифференцировке с утратой злокачественности. Что касается опухолей беспозвоночных и низших позвоночных, то они изначально более дифференцированы и менее злокачественны, чем у млекопитающих.
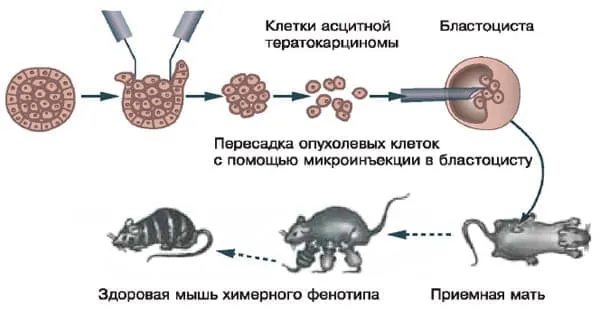
Опыты Минц и Ильмензее.
Пересадка опухолевых клеток в эмбрион здоровой мыши приводит к их трансдифференцировке в нормальные. При этом развиваются химерные мыши с характерной мозаичной окраской (мышь, у которой брали опухоль, была черной). Изображение: «Химия и жизнь»
Способность опухолевых клеток дифференцироваться в различных направлениях вместе с утратой злокачественности и активация в опухолях генов, которые не экспрессируются в нормальных тканях, но присутствуют в геноме, в том числе эволюционно новых генов, теоретически может приводить к возникновению эволюционно новых типов клеток.
Положительная роль патологий
До настоящего времени в науке господствует представление, что патологические процессы не могут играть положительную роль в эволюции. Но это не так. Наука знает примеры патологий и патогенов, имеющих приспособительное и/ или положительное эволюционное значение. Например, эволюции могут способствовать вирусы, переносящие гены между организмами разных видов.
Другой пример адаптивной молекулярной патологии — признак серповидноклеточности эритроцитов, связанный с некоторой устойчивостью к малярийному плазмодию Plazmodium falciparum. Эта болезнь вызвана альтернативной формой гемоглобина A, HbS. Эритроциты, содержащие такой гемоглобин, имеют серповидную форму, что приводит к повышению вязкости крови, замедлению кровотока и застою крови в капиллярах. Анемия, отечность, гемолиз, воспаление — вот неполный перечень симптомов серповидноклеточной анемии. Однако в районах с высоким распространением малярии частота мутантного аллеля HbS выше, поскольку гетерозиготные носители этого аллеля менее восприимчивы к малярии. Вообще, эволюция невозможна без мутационного процесса, хотя значительная часть мутаций (если не большинство) вредна для индивидуальных организмов.
Мутационный процесс имеет две стороны. Он приводит к нарушениям в сбалансированных молекулярных механизмах и работает как генератор разнообразных болезней, но он же поставляет новый генетический материал для отбора.
Все сказанное о мутациях справедливо и для опухолей. Хотя развитие опухоли вредно для индивидуальных организмов, но оно обеспечивает избыточные массы клеток с высоким уровнем биосинтеза, из которых в ходе эволюции иногда могут возникать новые клеточные типы.
Полезные опухоли
Бактерии рода Rhizobium вызывают на корнях бобовых опухоли — азотфиксирующие клубеньки. Населяющие их бактерии фиксируют азот, используемый самим растением, поэтому опухоль фактически превратилась в новый орган с регулируемой функцией. Существенное отличие клубеньков от обсуждавшихся выше опухолей, вызываемых A. tumefaciens, заключается в том, что опухоли, вызываемые агробактериями, полезны только самим бактериям, а растению вредны.
Для фиксации азота необходим леггемоглобин, который составляет около 40% растворимых белков клубеньков и кодируется геном растения. Ученые предполагают, что ген леггемоглобина — результат дупликации и последующей дивергенции гена, кодировавшего несимбиотический гемоглобин, и для бобовых относительно нов.
Следующий пример роли опухолей, реализовавшихся в эволюции, можно найти у рыб рода Xiphophorus, известных аквариумистам как меченосцы. По бокам тела и на спинном плавнике у них расположены темные маскировочные пятна, которые представляют собой гигантские меланоциты. В потомстве, полученном при скрещивании X. maculatus и X. helleri, у некоторых рыбок возникают меланомы. Феномен развития опухоли объяснили немецкие ученые Мулх Ахуджа и Фриц Андерс. Схема их эксперимента приведена на рисунке. Оказалось, что на половой хромосоме X. maculatus расположен «опухолевый локус» Tu, а на аутосоме — регуляторный локус R, подавляющий активность опухолевого гена. У X. helleri отсутствуют как Ти, так и R, поэтому при межвидовом скрещивании гибриды имеют только одну копию гена R, что приводит к усилению экспрессии генов Tu и увеличению размеров темных пятен на теле у рыб. А при скрещивании гибридных самок с самцами X. helleri у части потомков ген R вообще отсутствует, и при наличии локуса Tu развивается злокачественная меланома.
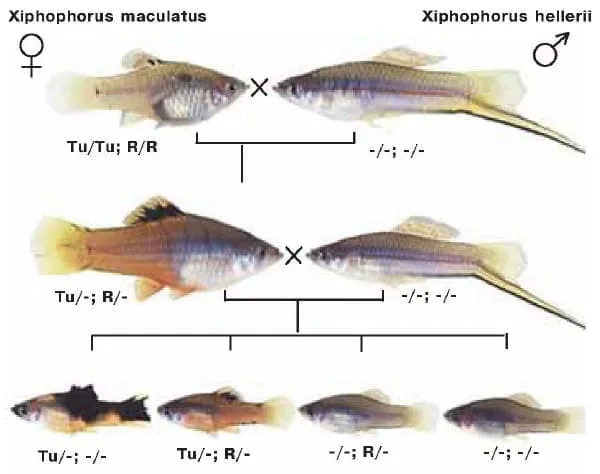
Меланома у меченосцев развивается только в том случае, если рыбка имеет хотя бы одну копию гена Tu, но ни одной копии гена R (пояснения в тексте). Поэтому при межвидовом скрещивании, показанном на схеме, в третьем поколении у четверти особей появляются меланомы. Изображение: «Химия и жизнь»
Мы предполагаем, что в эволюции сначала появилась популяция рыб с локусом Tu и высокой предрасположенностью к развитию меланом. После этого мог возникнуть ген R, который блокировал развитие меланом, и это в конечном счете привело к появлению полезного для рыбы признака — маскировочных пятен (макромеланофор).
Позднее оказалось, что локус Ти включает два тесно сцепленных гена: Mdl, определяющий различные типы пигментирования, которые связаны с макромеланофорами (Mdl -macromelanophore determining locus), и онкоген Xmrk. Оба гена относительно новы и возникли незадолго до появления рода Xiphophorus (5–6 миллионов лет назад). Самки Xiphophors maculatus предпочитают самцов с увеличенной экспрессией меланина, связанной с присутствием онкогена Xmrk. Макромеланофоры у X. maculatus представляют собой меланому, стабилизировавшуюся в результате удачной комбинации онкогена, гена-супрессора и определяющего пигментацию гена Mdl. Благодаря этой комбинации половой отбор поддерживает присутствие онкогена в популяции.
Мы можем также привести пример искусственного отбора доброкачественной опухоли у золотых рыбок Carassius auratus var. oranda. Их декоративная «шапочка» представляет собой папиллому (по личному сообщению М. А. Забежинского), но рыбки, обладающие этим признаком, успешно размножились благодаря искусственному отбору.
Нужны новые концепции!
Еще одной книгой, существенно повлиявшей на развитие моих представлений о возможной эволюционной роли опухолей, стала книга Сусуму Оно «Эволюция путем дупликации генов». Изложенные в ней представления о возникновении новых генов из избыточных копий предковых генов, ускользающих из-под контроля естественного отбора, до сих пор господствуют в теории молекулярной эволюции. А такими избыточными клеточными массами могут быть только опухоли.
В ходе эволюции генома постоянно возникают новые гены, но где им проявить себя? Уже имеющиеся ткани для этого не подходят. Число клеточных делений в этих тканях ограниченно, и они не могут снабжать эволюционирующие организмы избытком клеток, да и задачи у них другие. В дифференцированных тканях уже установился баланс между существующими генами, поэтому «новички» там обычно молчат. Для экспрессии «спящих» и/или эволюционно новых генов, возникавших в процессе эволюции генома в клетках зародышевой плазмы (но не в клетках опухолей), организму необходим некий «полигон», на роль которого идеально подходит опухоль.
В тех случаях, когда экспрессия нового гена в клетках опухоли приводила к возникновению новой функции, одновременно возникали новые обратные связи, регулирующие его деятельность. Клетки опухоли дифференцировались и давали начало новому для данного вида многоклеточных организмов типу клеток, который наследовался благодаря эпигеномным механизмам, как и предсуществовавшие типы клеток. Схема этого процесса показана на рисунке.
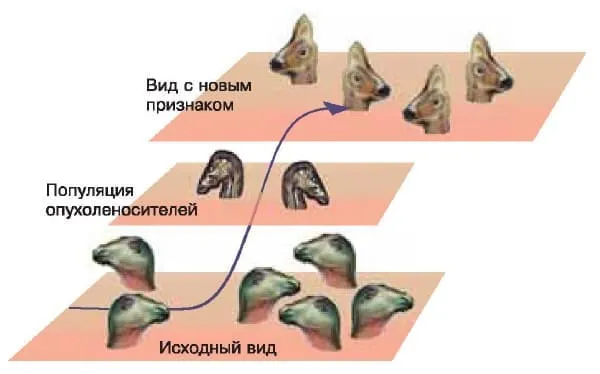
Популяция носителей опухоли — переходная форма между двумя видами, стоящими на разных ступенях прогрессивной эволюции. Изображение: «Химия и жизнь»
Популяции организмов-опухоленосителей, у которых опухоли были генетически или эпигенетически детерминированы, могли представлять собой переходные формы между видами организмов, находящихся на разных ступенях прогрессивной эволюции. Примером такого рода популяций опухоленосителей служат упомянутые выше гадрозавры.
В определенные периоды филогенеза дифференцировка опухолевых клеток в популяциях опухоленосителей должна была происходить достаточно часто, чтобы давать популяции организмов с новым типом клеток. Эти организмы затем проходили отбор на конкурентоспособность и приспособленность, как мы видим у рыб рода Xiphophorus.
Новые типы клеток могли принимать участие в образовании новых тканей и органов, например клубеньков бобовых или костных разрастаний на головах гадрозавров, с помощью которых они, как принято считать, издавали различные звуки.
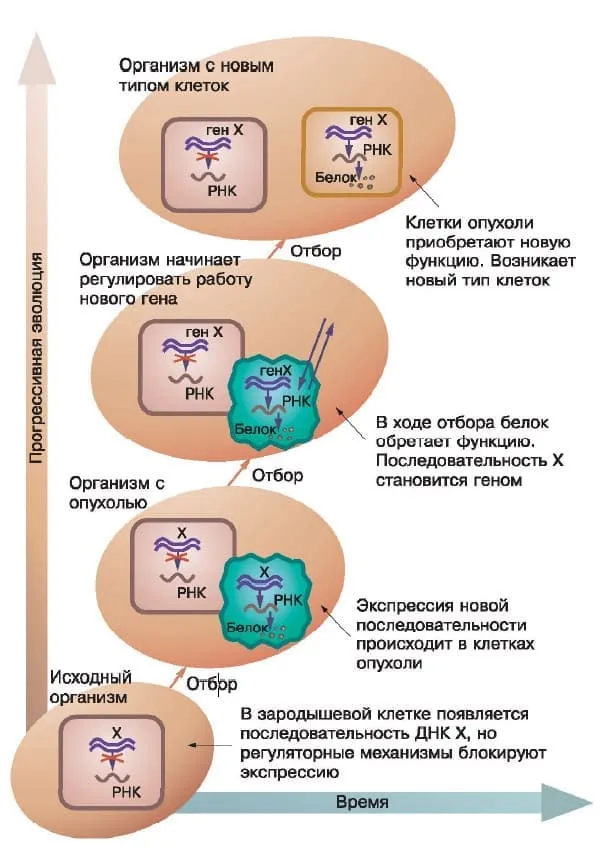
Возможно, опухоль — это колыбель нового типа клеток. Изображение: «Химия и жизнь»
У млекопитающих насчитывают от 200 до 500 специализированных типов клеток. Так что если опухоли действительно играли роль в происхождении новых клеточных типов, то происходило это достаточно редко.
Итак, в соответствии с концепцией эволюционной роли опухолей они предоставляют условия для экспрессии эволюционно новых и/или спящих генов, которые не экспрессируются в нормальных клетках и потому не подвержены в них действию естественного отбора и не имеют функции. Изменение этих последовательностей ДНК в зародышевых клетках и их экспрессия в опухолях могут привести к возникновению гена с новой функцией, что в свою очередь вызовет дифференцировку опухолевых клеток в соматические клетки нового типа. Очень важно то обстоятельство, что для возникновения нового гена и новой функции недостаточно только эволюции последовательности на уровне ДНК. Необходимы также ее экспрессия и отбор на новую функцию на уровне РНК и /или белка, который должен начаться в определенной момент эволюции новой последовательности.
Предсказания и их подтверждение
Концепция эволюционной роли опухолей позволяет выдвигать экспериментально проверяемые гипотезы. Так, мы предположили, что в опухолях должно экспрессироваться много последовательностей, которые не проявляют себя ни в каких нормальных типах клеток, причем последовательности эти либо эволюционно новые, либо ранее молчащие, и что в опухолевых клетках происходит отбор на новые функции. Начиная с 1980-х годов, мы работали над экспериментальным подтверждением этих гипотез и показали, что в опухолях млекопитающих экспрессируются последовательности, не экспрессирующиеся ни в одной экспериментально доступной нормальной ткани.
Но к концу 1990-х годов в базах данных появилось множество новых последовательностей ДНК из самых разнообразных нормальных и опухолевых тканей. Мы решили провести глобальное сравнение всех известных последовательностей, которые экспрессируются в опухолевых и нормальных тканях. По существу, мы повторяли эксперименты с «суммарными» препаратами РНК, но на более высоком методическом уровне.
Чтобы выявить гены, которые экспрессируются только в опухолевых тканях, мы провели компьютерный анализ последовательностей и действительно обнаружили десятки таких генов, то есть результаты были сходны с теми, которые мы получили ранее методом молекулярной гибридизации. Среди выявленных генов есть несколько уже известных опухолевых маркеров — это служит своеобразным внутренним контролем, подтверждающим, что компьютерный подход «работает». Около половины найденных последовательностей не имеют известной функции. Один из специфичных белков оказался перспективным иммуногеном для создания противоопухолевой вакцины. В дальнейшем мы неоднократно повторяли сравнение нормальных и опухолевых последовательностей, используя новые данные и усовершенствованные программы, просмотрели в итоге более 7 млн последовательностей и подтвердили полученные результаты.
Насколько эволюционно новы обнаруженные нами опухолеспецифические последовательности? Среди первых девяти изученных новыми оказались шесть: две из них возникают у человека, одна — у приматов, а три другие найдены только у млекопитающих и эволюционируют нейтрально, о чем свидетельствуют высокая скорость нуклеотидных замен и отсутствие консервативных фрагментов в их составе.
Затем мы применили противоположный подход — исследование опухолевой специфичности генов, о которых известно, что они эволюционно новые. В настоящее время, когда полностью секвенированы геномы многих видов организмов, такие гены можно обнаружить. И первый же изученный нами ген (PBOV1, prostate and breast cancer overexpressed 1), эволюционно новый для человека, оказался абсолютно опухолеспецифичным — он экспрессировался в 20 опухолях различной локализации и ни в одной из 19 нормальных тканей!
Таким образом, предположение об экспрессии эволюционно новых и/или спящих последовательностей в опухолях получило экспериментальное подтверждение.
Объяснение парадоксов
Совсем недавно ученые из разных зарубежных лабораторий получили данные о скорости эволюции некоторых опухолеспецифических генов у разных видов млекопитающих, включая приматов. Для этого последовательности опухолеспецифических генов человека сравнивали с последовательностями аналогичных генов приматов и мыши. Оказалось, что скорость эволюции таких генов выше, чем у нормальных тканеспецифических генов, что свидетельствует об отборе на новую функцию. Эти данные можно рассматривать как прямое свидетельство в пользу нашей концепции. Действительно, если эволюционно новый ген экспрессируется в опухоли или нейтральная последовательность приобретает функцию и становится эволюционно новым геном, должен иметь место отбор на усиление новой функции, который связан с более высокой скоростью замен в нуклеотидной последовательности. Именно это и было обнаружено в упоминавшихся работах. Следует подчеркнуть, что речь идет об эволюции организмов, а не об эволюции злокачественных клеток опухоли. Парадокс позитивной селекции опухолеспецифических генов в эволюции организмов нельзя объяснить иначе, как допустив позитивную роль опухолей в эволюции.
Рассмотренные выше бобово-ризобиальный симбиоз и маскировочные пятна у рыб рода Xiphophorus также связаны с экспрессией в опухолях эволюционно новых для организма генов, которая сопровождалась приобретением функции и утратой автономности опухолевыми клетками. Список подобных примеров можно продолжить.
Новые парадигмы — новые технологии
Итак, мы видим, что опухоли в ряде случаев могли играть роль в эволюции, снабжая многоклеточные организмы избыточными клеточными массами для экспрессии новых генов. Популяции организмов-опухоленосителей, возможно, служили переходными формами между разными видами, которые отличаются количеством типов клеток.
Концепция возможной эволюционной роли опухолей позволяет объяснить механизмы возникновения новых типов клеток у многоклеточных, что до последнего времени упрощенно сводили только к появлению новых генов. Она также объясняет парадокс позитивной селекции множества опухолеспецифических генов в ряду млекопитающих и приматов, необъяснимый другим путем. Она позволяет формулировать нетривиальные предсказания, планировать эксперименты по их подтверждению и получать оригинальные результаты.
Концепция эволюционной роли опухолей, как всякая новая парадигма, значительно расширяет наши представления о природе опухолей и о возможностях влияния на опухолевые процессы. Из нее следует, что в области лечения и профилактики онкологических заболеваний следует больше внимания уделять подходам, связанным с возможностью дифференцировки опухолевых клеток и их включения в функциональные сети организма; что следует искать эволюционно новые опухолеспецифические гены как возможные мишени терапевтических и/или профилактических воздействий, а также использовать их в молекулярной диагностике.

Для комментирования материалов регистрация на сайте не нужна. Комментарии проходят премодерацию. Email комментаторов не публикуется.