
Ионные каналы, гены и эволюция

В. В. Зюганов, Ю. А. Лабас, В. В. Хлебович
Журнал «Природа» № 91987
Экологические последствия научно-технической революции привлекают внимание не только практиков, технологов и специалистов по охране природы. Эти последствия заставляют повернуться к себе лицом таких, казалось бы, чистых теоретиков, как специалисты по теории естественного отбора.
Десятилетиями идут споры о его действенности в быстро меняющихся условиях внешней среды. Противники дарвиновской теории эволюции утверждают, что в короткие сроки неопределенная изменчивость (мутации) как материал для естественного отбора не может привести к приспособлению видов к новой среде обитания, договариваются до невозможности происхождения человека от примитивной клетки за историю жизни на Земле.
Однако прямые эксперименты поставляют доказательства для подтверждения дарвиновских идей, позволяют уже не дедуктивно, а индуктивно подойти к решению острейшей проблемы дарвинизме, делают пресловутую «непрактичность» теорий действенным орудием практичности. Изменение важнейшего экологического свойства — смена пресных вод на соленые — оказывается следствием мутации нескольких копий одного лишь гена. Для таких мутаций не нужен длительный эволюционный период, они позволяют организму быстро приспособиться к жизни в совершенно непривычной среде. Об экспериментально добытых фактах эволюционного значения, касающихся изменения ионной проницаемости клеточных мембран, и пойдет речь.
Об авторах публикации

Валерий Валериевич Зюганов, кандидат биологических наук, научный сотрудник лаборатории постнатального онтогенеза Института биологии развития им. Н. К. Кольцова АН СССР. Занимается генетикой, экологией, систематикой и эволюцией рыб.

Юлий Александрович Лабас, кандидат биологических наук, старший научный сотрудник лаборатории поведения низших позвоночных Института эволюционной морфологии и экологии животных им. А. Н. Северцова АН СССР. Область научных интересов — биолюминесценция, физиология, онтогенез клетки.

Владислав Вилгельмович Хлебович, доктор биологических наук, профессор, специалист по зоологии беспозвоночных, главный научный сотрудник Зоологического институте АН СССР. Основные научные труды посвящены физиологии, механизмам адаптации и эволюции животных, биологии эстуариев, систематике и экологии кольчатых червой. Автор монографий: Критическая соленость биологических процессов. Л., 1974; Акклиматизации животных организмов. Л., 1981.
Путешествие ионов
Любая клетка, живет ли она свободно или входит в состав многоклеточного организма, контактирует со сложными растворами различных неорганических ионов. Чаще всего это вода, пресная или морская, а для клеток многоклеточного организма — различные внутренние среды, в том числе плазма крови. Сама же клетка, отделенная от «окружающей ее среды клеточной (плазматической) мембраной, имеет свой собственный набор ионов в цитоплазме. Непременное условие сохранения жизнеспособности клетки — асимметричное распределение ионов по обе стороны ее мембраны, постоянный обмен ионами между вне- и внутриклеточной средами.
Клеточная мембрана, как известно, — это двойной слой липидных молекул, в который встроены различные белки. Перекос ионов непосредственно сквозь липидные слои невозможен, он осуществляется только с участием мембранных белков. В настоящее время различают три способа переноса ионов через клеточную мембрану: движение по каналам, обмен одних ионов на другие посредством особых переносчиков и активный транспорт.
Движение ионов по каналам и с переносчиками, в отличие от активного транспорта, происходит пассивно, т. е. по направлению концентрационного и электрохимического градиентов — от большего к меньшему. Для каждого из основных ионов естественной среды — натрия, калия, кальция, хлора и, по-видимому, магния — осуществляют специализированные каналы. Кроме того, ионные каналы подразделяются на невозбудимые (постоянно открытые) и возбудимые, которые активируются (открываются) только временно, в ответ на тот или иной, специфичный для данного типа каналов, внешний стимул — механическое раздражение, химическое соединение, электрическое поле.
Роль обмена ионами с внешней средой в жизни клеток и организмов огромна. Обмен необходим нервным, мышечным и другим возбудимым клеткам, чтобы в них генерировались электрические импульсы, или так называемые потенциалы действия. Для этого в клетках раскрываются электровозбудимые каналы, через которые катионы натрия или кальция проникают из среды в цитоплазму, где их намного меньше, чем в среде, а затем раскрываются калиевые каналы, но через них катион калия выходит из цитоплазмы, где его больше, чем во внешней среде. Такое путешествие ионов приводит сначала к быстрому падению электроотрицательного потенциала, нередко он переходит через ноль и изменяет знак, а затем снова восстанавливается до исходного значения. Так в клетке возникают электрические импульсы.
Посредством переносчиков ионы движутся через клеточную мембрану опять-таки пассивно, но в обмен на другие ионы, например натрий часто обменивается на кальций или ион водорода (протон).
Активно транспортируются ионы через плазматическую мембрану против их концентрационного и электрохимического градиентов, т. е. оттуда, где этих ионов сравнительно мало, туда, где их больше. На такую работу расходуется энергия, которая освобождается при ферментативном гидролизе молекул аденозинтрифосфорной кислоты (АТФ). В частности, в плазматической мембране фактически всех клеток присутствует Na+ -К+ -АТФаза, фермент, который гидролизует АТФ и при этом откачивает ионы натрия из цитоплазмы в окружающую среду в обмен на ионы калия, переносимые в цитоплазму.
Повышенная концентрация ионов калия в цитоплазме в сочетании со сравнительно высокой его проницаемостью через плазматическую мембрану в покое — обычно основная причина электроотрицательного трансмембранного потенциала, достигающего у большинства клеток от -60 до -80 мв. И ионные каналы, и ферменты, ответственные за активный транспорт ионов, и, по-видимому, переносчики представляют собой белковые молекулы или же небольшие их комплексы, встроенные в липидный слой мембраны, т. е. образования, по сути, относительно простые. Так, уже упомянутый механизм встречного активного транспорта ионов натрия и калия, «натриевый насос», создается деятельностью всего одного фермента, или, иными словами, одного белка — Na+ -К+ -АТФезы. По новейшим данным, и электровозбудимый натриевый канал позвоночных животных тоже образован только одной молекулой гликопротеина с молекулярной массой около 260 кД. Правомерно полагать, что невозбудимые натриевые каналы организованы, по крайней мере, не сложнее возбудимых и представляют собой тоже отдельные молекулы белка, встроенные в липидный матрикс мембраны. Это же предположение может быть высказано относительно других ионных каналов, переносчиков и ферментов, которые осуществляют активный транспорт ионов через мембрану.
Есть данные, что для плазматической мембраны клеток любого типа характерна своя врожденная специфика качественного состава, свойств и количества ионных каналов, переносчиков и ферментов, перекачивающих ионы. Подобно любому другому наследственному признаку, гены, ответственные за синтез этих трех типов белков, по-видимому, могут подвергаться скачкообразным наследственным изменениям — мутациям. Но если каждый из типов переноса ионов обеспечивается только одним белком, достаточно всего одной мутации — мутации того единственного гена, в котором закодирован данный белок (канал, переносчик или транспортирующая АТФаза), чтобы ощутимо изменилась и наследственно закрепилась способность плазматической мембраны пропускать сквозь себя или же активно перекачивать тот или иной ион. Считается, что такие мутации могут служить предпосылкой для относительно быстрых эволюционных перестроек, поскольку функционально важный признак контролируется всего одним геном, а не сложным их комплексом. Вместе с тем имеются веские основания полагать, что мутации, обеспечивающие измененную ионную проницаемость клеточной мембраны, чреваты последствиями в целом ряде аспектов жизнедеятельности организмов, в том числе в их поведении и выживании в средах разной солености, в переселении из пресных вод в морские, в освоении засоленных почв, в приспособлении к недостатку влаги в пустынях и т. д.
Нам известны две группы фактов, непосредственно подтверждающих эти предположения.
«Пешки», «параноики» и другие
Хорошо знакомая всем со школьных времен инфузория парамеция, или туфелька, характеризуется быстрым маневренным движением. Оно создается ритмическим биением ресничек, покрывающих поверхность этого одноклеточного животного. Оказалось, что с помощью химических мутагенов можно изменить обычную манеру парамеции двигаться.
Американский исследователь А. С. Кунг с сотрудниками посредством одного из таких мутагенов (М-метил-N-нитро-N-нитрозогуанидина) получил более 15 типов мутантов парамеций с явно выраженными аномалиями их движений. В электрофизиологических экспериментах и в опытах по реактивации ресничного движения после разрушения клеточных мембран мылоподобными веществами (детергентами) он доказал, что причина большинства этих аномалий — отклонения от нормы свойств возбудимых кальциевых или калиевых каналов или их количества. Заметим, что возбудимые натриевые каналы, чувствительные к специальному блокирующему веществу — тетродотоксину, обнаружены пока только в мембранах нервных и других клеток высших беспозвоночных и позвоночных животных. В генетических опытах со скрещиванием выяснилось, что многие варианты аномального движения парамеций определяются мутацией всего одного аллельного гена. Некоторые мутанты Кунга теряли весьма типичную для ресничных инфузорий способность двигаться «задом наперед», поскольку направление, в котором бьются реснички, изменилось на 180°. У нормальных особей эти изменения возникают при столкновении переднего конца клетки с механическим препятствием (от которого парамеция стремится уйти) или в присутствии химических соединений, которых она избегает (например, хлористого натрия). Мутанты, неспособные двигаться задним ходом, при любых обстоятельствах вынуждены двигаться только вперед, подобно шахматным пешкам, за что и получили прозвище «пешки».
С помощью электрофизиологических методов Кунг показал, что причина необычного движения «пешек» кроется в отсутствии в их плазматической мембране электровозбудимых кальциевых каналов. Посредством скрещивания Кунгу удалось выяснить, что эти каналы исчезли из-за рецессивной мутации в каком-то одном из пяти несцепленных между собой генов. У нормальных инфузорий обратное биение ресничек вызывается активацией (приоткрыванием) механочувстеительных или хемочувствительных кальциевых каналов. Такие каналы, однако, сами по себе вызывают лишь ту минимальную деполяризацию клетки, которая требуется для последующей активации гораздо более многочисленных электровозбудимых кальциевых каналов.
При их отсутствии у «пешек» в цитоплазму инфузорий из окружающей пресной воды быстро проникает по концентрационному и электрохимическому градиентам весьма значительное количество ионов кальция (в покое его внутриклеточная концентрация обычно не более Ю-7 М, т. е. приблизительно в 104 раз ниже концентрации Са2+ в пресной воде и в 106 раз — в морской; при возбуждении клетки эта концентрация может быстро возрасти в 10–100 раз). В клетке кальций соединяется с находящимся в основании ресничек особым кальций-связывающим белком — чальмодулином, что приводит к их переориентации и обратному биению. Почему кальмодулин, связанный с ионами кальция, оказывает такое действие — пока неясно.
У другого мутанта парамеции, прозванного «параноиком», движение задним ходом, обусловленное обратным биением ресничек, гипертрофировано. Оно не только затягивается каждый раз, когда инфузория натыкается на механическое препятствие или ощущает неприятное для нее вещество, но и часто возникает без видимых внешних воздействий на клетку. Причин, по-видимому, две: одна — замедленное закрывание электровозбудимых кальциеаых каналов, вторая — намного превышающая норму чувствительность к присутствующим в воде ионам натрия, которые вызывают у парамеции реакцию избегания. Эти «параноики» — последствие единственной доминантной мутации опять-таки в одном из пяти несцепленных генов.
Еще два мутантных варианта, названных Кунгом «быстрые-1» и «быстрые-2», — это парамеции, очень быстро плавающие за счет аномально частого биения ресничек. Быстрые туфельки могли появиться из-за увеличенного против нормы количества калиевых каналов в плазматической мембране, приводящего к ее сверхполяризации — аномально высокому трансмембранному потенциалу. Высокий потенциал, в свою очередь, может вызывать усиленный поток ионов кальция, поступающих через невозбудимые каналы в цитоплазму по электрохимическому градиенту. Такое усиление, в отличие от аналогичного, но большего, связанного с активацией электровозбудимых каналов, не изменяет направление ресничных биений, но ускоряет их.
Кунгом описаны также мутанты парамеции, которые превращаются в «пешек» каждый раз, когда температура воды превышает 30° С, а при более низкой они ведут себя нормально.
Мутации, выявленные Кунгом, проявляются прежде всего в аномальном движении клеток. Самое главное в его экспериментах — доказательство того, что из-за значительных изменений ионной проницаемости клеточной мембраны, обусловленных мутацией всего одного гена, могут меняться жизненно важные функции организма. По-видимому, не имеет решающего значения, происходит это в одно- или многоклеточном организме. Вполне вероятно, что аналогичные наследственные изменения ионной проницаемости клеточных мембран способны возникать в разных органах и тканях и у многоклеточных организмов, делаясь причиной генетических аномалий нервной деятельности, функций выделительных органов и т. д. Не исключено, что некоторые из таких или подобных им мутаций имеют и экологическое значение, поскольку изменяют отношение клеток к солености внешней среды. В частности, у водных организмов подобного рода изменения могут содействовать или, наоборот, препятствовать их прорыву через экологический барьер море — пресная вода.
Загадки трехиглой колюшки
Вторая группа фактов, действительно подтверждающих связь мутаций ионной проницаемости клеточных мембран с экологическими возможностями организма, выявлена авторами этой статьи.
У трехиглой колюшки (Gasierosteus aculeatus) описано три формы наследственной внутривидовой изменчивости: форма trachurus, форма leiurus и форма semiarmatus. Все они свободно скрещиваются между собой, но заметно различаются как внешне (по характеру покрытия тела типичными для колюшек костными щитками), так и экологически — отношением к солености среды. У формы trachurus костные щитки тянутся вдоль всего тела сплошной полосой от головы до основания хвоста; колюшка этой формы может жить и размножаться и в пресной, и в морской воде. В бассейне Белого моря она образует и морские, и проходные, и пресноводные оседлые популяции, но на Дальнем Востоке, в США, Канаде ведет себя только как проходная рыба: живет в море, а размножается в пресных водах. Интересно, что сперматозоиды формы trachurus приобретают способность двигаться в пресной воде только после нескольких часов пребывания в ней самца.
У формы leiurus костные щитки покрывают только переднюю часть тела до уровня грудных плавников. Эта форма обитает исключительно в пресных водоемах, хотя во взрослом состоянии может неограниченно долго жить и в морской воде, если ее температура превышает 4° С. Оплодотворение яйцеклетки и дальнейшее развитие икры у формы leiurus тоже вполне возможны в морской воде, тех же успехов можно добиться и в экспериментах, если 2 раза в сутки менять пресную воду на морскую, т. е. имитировать приливно-отливный ритм, обычный в наших северных морях — Белом и Баренцевом. В то же время сперматозоиды формы leiurus, в отличие от сперматозоидов формы trachurus, неподвижны в морской воде, что и служит, по-видимому, главным препятствием переселения leiurus из пресной воды в морскую.
Форма semiarmatus — гибрид двух упомянутых форм. Она занимает промежуточное положение между ними и по характеру покрытия тела щитками, и отношением к солености воды.
Сперматозоиды рыб, как и подавляющего большинства других организмов, в состоянии оплодотворить яйцеклетку, только если они активно движутся к ней, привлекаемые специально испускаемым этой клеткой веществом. Их движение создается биением жгутиков, образований очень близких к ресничкам инфузории парамеции. Благодаря тем или иным особенностям концентрации или ионного состава семенной жидкости, сперматозоиды в ней, как правило, неподвижны. Этим, очевидно, предотвращается бессмысленная растрата ими энергии до ситуации, ведущей к оплодотворению. Попав в среду, в которой нормально происходит оплодотворение (у организмов с наружным оплодотворением — а воду), сперматозоиды сперва активно плавают, отыскивая яйцеклетку, а затем, если оплодотворение не произошло, еще некоторое время «дрожат» на месте и погибают. Устойчивость сперматозоидов к факторам среды обычно оценивают по длительности их активного плавания — поступательного движения. Воспользовавшись этим показателем, мы сравнили солевую устойчивость сперматозоидов трех форм трехиглой колюшки из бассейна Белого моря.
Выяснилось, что мужские половые клетки пресноводной формы leiurus теряют способность двигаться при солености среды 18% (соленость вод Белого моря около 25%). Сперматозоиды морской формы trachurus становятся неподвижными только при 52%. Что же мешает сперматозоидам пресноводной формы активироваться при высокой солености воды?
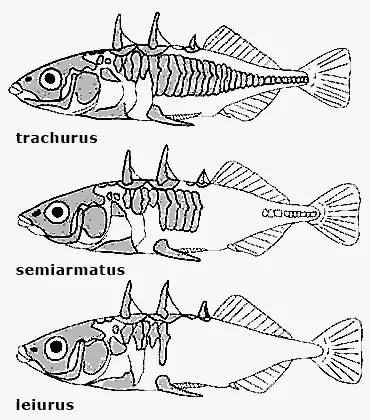
Формы трехиглой колюшки.
Мы проверили, не связано ли обездвиживающее действие морской воды на сперматозоиды формы leiurus с каким-то одним из основных, входящих в состав воды катионов (Na+, К+, Са2+ и Mg2+) или анионов (Cl-, SO2-4). Для этого мы сравнили движение сперматозоидов в растворах чистых солей — хлоридов (NaCI, KCl, CaCI2 и MgCI2), а также сульфатов натрия и калия (Na2SO4 и K2SO4), приготовив эти растворы той же концентрации, в которой соли содержатся в морской воде. Кроме того, мы проверили, как влияет на движение тех же клеток в морской воде блокатор хлорных каналов плазматических мембран (возбудимых и невозбудимых) — фуросемид. Выяснилось, что специфическое обездвиживающее действие на сперматозоиды пресноводной формы колюшки оказывают только катионы натрия (в растворах NaCI и Na2SO4), независимо от того, хлорид или сульфат был анионом. Кстати, фуросемид видимых эффектов не вызывал.
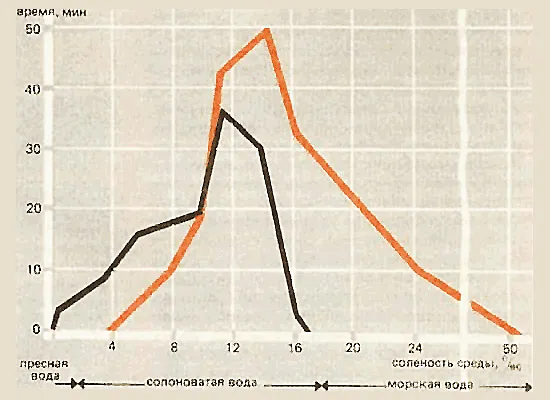
Продолжительность движения сперматозоидов пресноводной (черная кривая) и морской форм колюшки о иоде разной солености.
Далее мы попытались проверить, не различаются ли пресноводная и морская формы колюшки активностью ферменга Na+-К+-АТФазы. Вспомним, что этот фермент работает как насос при активном переносе через мембрану ионов натрия и калия против градиента концентрации. Оказалось, что и строфантин К, яд, угнетающий этот фермент, не оказывает специфического влияния на какой-то один из сравниваемых генотипов, а высокой концентрации он только тормозит движение сперматозоидов любой формы колюшки. Естественно, не стимулировал движение сперматозоидов пресноводной колюшки и субстрат АТФазы — аденозинтрифосфорная кислота (если бы различия в подвижности сперматозоидов были связаны с недостатком субстрата, его добавление в среду вызывало бы активацию половых клеток).
Нам оставалось предположить, что инактивация сперматозоидов пресноводной формы колюшки в морской воде связана с пассивной диффузией ионов натрия внутрь клетки через каналы или переносчики мембран. Для проверки этого предположения в экспериментах мы располагали амилоридом, соединением, которое блокирует (затыкает) невозбудимые натриевые каналы клеточной мембраны.
Результат превзошел все наши ожидания. Добавленный в морскую воду в концентрации 10-5 М амилорид активировал движение сперматозоидов пресноводных форм колюшки. Блокатор натриевых каналов сделал эти сперматозоиды почти столь же выносливыми к высокой солености среды, как и половые клетки морской формы!
Остается предположить, что плазматическая мембрана сперматозоидов пресноводной и морской форм различается всего лишь количеством невозбудимых натриевых каналов на единицу площади — у пресноводной формы их должно быть больше. Может быть и другая причина: каналы почему-то начинают лучше пропускать ионы натрия. Результат действия и той и другой причины одинаков: избыток проникающих в сперматозоид катионов натрия угнетает движение жгутика. Скорее всего, в этом сказывается не прямое действие избытка катионов натрия, а связанное с ним последующее понижение цитоплазматической концентрации катионов кальция.
Известно, что из всех катионов морской воды только кальций и магний необходимы для активации жгутиков мужских половых клеток, но цитоплазматическая концентрация ионов магния, хотя и относительно велика, мало меняется. Ионы кальция действуют через упомянутый уже кальций-связывающий белок (кальмодулин), который активирует фермент аденилатциклазу, что ведет к образованию циклического адеиозинмонофосфата (цАМФ), вещества-посредника, активирующего еще один фермент — протеинкиназу. В результате цепочки этих реакций к особому белку «15К» (аксокинину) в структуре жгутика присоединяется неорганический фосфат и жгутик начинает активно двигаться. В то же время в плазматической мембране многих клеток (можно полагать, и сперматозоидов пресноводных форм колюшки тоже) присутствует натрий-кальциевый переносчик, благодаря которому из клетки в воду поступают ионы кальция в обмен на входящие в нее ионы натрия. При их высокой концентрации в среде клеткам с таким переносчиком в мембране угрожает значительное понижение цитоплазматической концентрации кальция. Возможно, что наличие в мембране еще одного переносчика — найтрий-протонного — может приводить к защелачиванию цитоплазмы, а это тоже неблагоприятно сказывается на сперматозоидах.
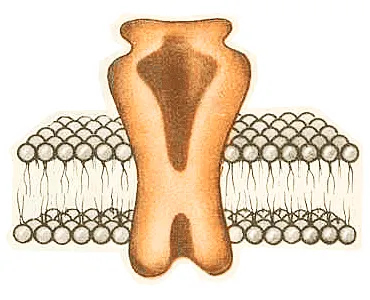
Модель трехмерной структуры натриевого канала в двуслойной мембране клетки. [По: Овчинников Ю. А. и др.//Биол. мембраны. 1985. Т. 2. № 10. С.060.]
Косвенным подтверждением того, что ионы натрия обездвиживают мужские половые клетки пресноводной формы колюшки, способствуя понижению цитоплазматического содержания ионов кальция, служит эффект среды, в которой была повышена его концентрация и концентрация верапамила — вещества, блокирующего кальциевые каналы мембран. Морская вода с повышенным в 5 раз содержанием кальция продлевала движение сперматозоидов, а верапамил не только обездвиживал их, но и снимал защитное действие на них амилорида у формы leiurus в морской воде.
Следует отметить, что обездвиживающее действие ионов натрия на сперматозоиды пресноводной колюшки напоминает аналогичное действие калиевых ионов на сперматозоиды лососевых рыб. Японский исследователь М. Морисава установил, что у этих рыб семенная жидкость характеризуется более высоким, в сравнении с пресной водой, содержанием ионов калия, который, по-видимому, деполяризует мембрану мужских половых клеток и тем самым замедляет вход в них ионов кальция через невозбудимые каналы по электрохимическому градиенту (сравним причины учащенного биения ресничек у «быстрых» мутантов парамеции). Морисава показал также, что сперматозоиды некоторых других пресноводных и морских костистых рыб обездвиживают отнюдь не ионы калия, а высокое (у пресноводных форм) или низкое (у морских) осмотическое давление семенной жидкости, безотносительно к ее собственному солевому составу.
Сравнительный анализ причин неподвижности сперматозоидов в семенной жидкости морской и пресноводной форм колюшки пока не проведен. Весьма вероятно, что у пресноводной формы тормозящую функцию выполняет именно ион натрия.
Таким образом, существенные экологические различия по столь важному для водных организмов признаку, как отношение к солености, у сперматозоидов морской и пресноводной форм колюшки определяются различиями в количестве или устройстве натриевых каналов клеточной мембраны. Каждый такой канал сформирован, скорее всего, одной молекулой белка. Поэтому соответствующие отличия должны, по-видимому, определяться мутациями одного гена, хотя бы и представленного несколькими копиями, как у парамеций.
Чтобы установить, сколько копий гена обеспечивают различия в солеустойчивости сперматозоидов разных форм колюшки, мы провели опыты по скрещиванию форм trachurus и leiurus. Получив 8 поколений потомства, мы проанализировали распределение признака солеустойчивости сперматозоидов 103 самцов и сравнили полученные результаты с теоретически ожидаемым расщеплением (полагая, что признак солеустойчивости определяется пятью копиями двухаллельного гена, именно для этого случая и был произведен расчет). Оказалось, что доля солеустойчивых спермиев в потомстве различается у разных самцов, но всегда кратна 1 /8, или 12,5%, такой же она должна быть и по теоретическим расчетам. Совпадало и число фенотипических классов: и по теории, и у реальных потомков их было 9.
Следовательно, солеустойчивость сперматозоидов колюшки обеспечивается пятью идентичными генами, по два аллеля в каждом. Любое другое количество копий дает высокодостоверные различия между наблюдаемыми и ожидаемыми частотами встречаемости признака в поколениях. Устойчивость к солям, по-видимому, возникает только в тех случаях, когда мутантными оказываются 3 и более из этих 5 генов, т. е. признак ведет себя как количественный.
От проницаемости мембран к эволюционным преобразованиям
Итак, есть все основания полагать, что и у парамеции, и у трехиглой колюшки — организмов, принадлежащих к разным типам животного мира, важнейшие экологические параметры и характеристики поведения могут изменяться в результате мутации всего лишь одного гена, пусть даже и представленного несколькими копиями в разных хромосомах. У парамеций большое количество различных мутаций удалось получить искусственно за короткий срок. Попыток вызвать мутации, которые изменяли бы отношение организма к таким факторам внешней среды, как соленость, действуя на животных химическими мутагенами, пока сделано не было. Но перспективность таких работ нам кажется несомненной.
Индуцирование мутаций солевой устойчивости половых клеток или клеток раннего зародыша пресноводных и в особенности проходных рыб (к ним относятся самые ценные промысловые рыбы — осетровые и лососевые), могло бы иметь важное хозяйственное значение, поскольку соленость многих пресных водоемов СССР неуклонно повышается. В таких условиях инкубация развивающейся икры ценных видов в солоноватой или морской воде была бы вполне перспективной.
В начале статьи мы говорили, что данные о мутациях ионной проницаемости мембран позволяют не только понять, за счет чего могут радикально изменяться экологические характеристики организма. Результаты наших экспериментов прямо подтверждают идеи дарвинизма. Ведь один из основных аргументов противников дарвиновской теории эволюции часто сводится к тому, что реальная эволюция протекает слишком быстро в сравнении с таким, якобы, невероятно медленным процессом, как возникновение неопределенной изменчивости в качестве материала для естественного отбора. Мутанты Кинга и выявленные нами причины, изменяющие ионную проницаемость мембран, могут служить наглядными примерами для иллюстрации несостоятельности этого аргумента. Складывается впечатление, что его сторонники исходят из представлений только о полигенном характере большинства адаптивных признаков, обеспечивающих организмам возможность внедряться в новую для них среду (экологическую нишу).
Что это представление ошибочно, свидетельствуют не только частные случаи с парамецией и трехиглой колюшкой, но и удивительно быстрое приспособление некоторых организмов к экологическим последствиям научно-технической революции — урбанизации, разнообразным загрязнениям среды (пестицидам, антибиотикам и т. д.). Это приспособление, несомненно, основывалось на дарвиновском принципе неопределенной изменчивости и отбора, несмотря на свой сверхбыстрый темп. Однако оно, как мы полагаем, осуществлялось в основном за счет мутаций минимального числа генов, возможно — единственного гена.
Неудивительны поэтому большие экологические различия, иногда присущие близкородственным видам, относящимся к одному роду. Так, например, среди инфузорий рода Euplotes и раков-бокоплавов рода Gammarus описаны виды и морские, и пресноводные. Едва ли такая экологическая специфика определялась большим числом генов, а не каким-то одним геном или же малым их числом. Ведь мутация всего одного регуляторного гена может повлечь за собой переход из функционального состояния в неактивное («спящее») или, наоборот, привести к экспрессии (при определенных ситуациях: стадиях развития, изменениях среды и т. п.) сразу большого комплекса структурных генов, а значит, и к проявлению нового свойства в фенотипе.
Нам пока неизвестны тонкости генетических превращений, которые изменяют солевую устойчивость трехиглой колюшки. Несомненно, однако, что частный случай — изменение проницаемости клеточных мембран — связан с далеко идущими последствиями в размножении и экологии организмов, а значит, и с их эволюцией.
Скачать в формате .pdf

Для комментирования материалов регистрация на сайте не нужна. Комментарии проходят премодерацию. Email комментаторов не публикуется.